Des bactéries et des fourmis
Alain Lenoir mis à jour 06-Fév-2026
Les bactéries des fourmis sont connues depuis longtemps mais on s'intéresse de plus en plus à elles depuis le développement des résistances aux antibiotiques. On trouve des bactéries symbiotiques dans l'intestin, elles aident au métabolisme et à l'immunité de la fourmi. On en trouve aussi sur la cuticule et partout sur et dans le corps de la fourmi. Elles forment le microbiome comme chez tous les animaux y compris l'homme. Ce microbiome participe à de très nombreux aspects de la biologie de la fourmi et de toute la vie de la société, par exemple lors des échanges trophallactiques et des léchages interindividuels, et même dans la formation de l'odeur coloniale. Elles participent aussi à la mémorisation chez les bourdons. C'est aussi un bel exemple d'évolution pour la symbiose. On parle maintenant d'holobionte.
Les
idées sur l'évolution sont en pleine mutation, en effet tout individu
(animal, plante) est en fait un véritable écosystème
et il comprend toute une série d'organismes symbiotiques comme des bactéries,
champignons et micro-algues et l'évolution agit sur cet ensemble.
A voir le très beau documentaire d'Arte : Ces
microbes qui nous gouvernent.
On y parle de l'homme bien sûr, des hyènes, des
souris, de guêpes, de seiches... Les billets de banque sont toujours "sales"
selon Barthélémy (2016) puisqu'ils portent de très nombreuses
bactéries. Les activités humaines transforment radicalement la
circulation des micro-organismes (bactéries,
champignons, micro-algues) sur la planète, avec des répercussions
probables sur les écosystèmes et les populations. On
trouve par exemple une dispersion considérable de l'interféron
de classe 1 qui favorise la résistance aux antibiotiques (Zhu
et al 2017, voir Luk 2018). La médecine légale s'intéresse
de plus en plus aux bactéries qui signent le passage d'une personne (Cabut
et Herzberg 2017). Wilson,
dans son livre de souvenirs en 1994 parlait déjà de la formidable
biodiversité des bactéries (pdf).
Pourtant, on va de plus en plus faire face aux bactéries
résistantes aux antibiotiques. Par exemple, en Inde où l'on fabrique
la plupart des antibiotiques utilisés, de nombreuses résistances
apparaissent, ce qui risque de poser des problèmes au niveau mondial
(Barnéoud et Bomboy 2018). Le
microbionte des fourmis intéresse de plus en plus de monde. Un article
de chercheurs du Costa Rica fait le bilan pour les Camponotus (citant
nos travaux sur les Blochmania pour l'immunité), les Cephalotini
et les Attini, essentiellement pour les symbioses nutritives. C'est une source
nouvelle pour la recherche d'antibiotiques (Artavia-León et al 2019).
Pascal Picq (2013) dit : « Ainsi, les pires atteintes à la
biodiversité susceptibles de mettre en danger l’avenir de l’humanité
ne viendront pas de l’extinction des tigres ou des gorilles, mais de la
disparition de ces multitudes de micro-organismes qui sont des partenaires silencieux
de notre évolution. La course à l’asepsie est devenue une
ineptie anti-évolutionniste dont les maladies nosocomiales ne donnent
qu’un petit aperçu. Réapprenons à vivre avec les
micro-organismes ! »
Science & Vie de janvier
2021 présente une revue complète sur le microbiome qui intervient
chez l'homme dans des domaines de plus en plus nombreux comme les relations
sociales (par exemple aussi chez les drosophiles), le développement,
l'immunité, les gouts, les émotions. Le corps humain est une véritable
symbiose avec des milliards d'organismes microscopiques (y compris des champignons).
En ce qui concerne la vie sur terre, "L'interdépendance est la règle"
entre tous les organismes et leur microbiote (par exemple les abeilles - Vernier
et al 2020). On parle
maintenant d'holobionte et d'hologénome
qui rassemble microbiotes et l'hôte.
Le microbiome devient un sujet
très important. Voir par exemple le dernier numéro de Science
et Vie. Les insectes sociaux bien sûr sont concernés. Sur Google
Scholar, en 2020 on trouve 1 050 citations avec la référence "microbiome
ants" contre 50 en 2010...
Hologénome : un nouveau nom qui apparait dans la littérature. Dans Bibliovie, il n'existe pas avant 2000 et 91 fois pour Google Scholar, 216 fois pour la période 200/2019, 264 fois par an entre 2010 et 2019, puis explosion en 2020 avec 594 fois.
Voir "Le charme discret de l'intestin" de Giulia Enders. On découvre tous les jours un peu plus le rôle des microbes. J'ai commencé à m'y intéresser quand, avec mon étudiant Danival Souza, on a découvert des bactéries symbiotiques chez les fourmis Camponotus. Depuis on s'aperçoit que les microbes font partie de notre organisme. Il faut repenser l'évolution non pas en terme d'individu mais d'écosystème complexe : il y a création d’entités entre un hôte et ses bactéries que l'on peut assimiler à un nouvel organisme, un superorganisme, ce que l'on peut qualifier de transition majeure (Kiers and West 2015). On a même créé la théorie de l'hologénome qui me semble excellente (Zilberg-Rosenberg and Rosenberg 2008). Giulia Enders reprend cette idée en disant "Depuis peu, la science considère l'être humain comme un véritable écosystème" (Enders p. 191).
De nombreuses espèces
de bactéries sont symbiotes d'insectes. Chez les abeilles, fourmis et
guêpes on trouve 197 espèces d'actinobactéries symbiotes,
essentiellement des Streptomyces et quelques Pseudonocardia
et Amycolatopsis (Matarrita-Carranza et al 2017). Le glyphosate pourrait
être une cause possible supplémentaire du déclin des abeilles
domestiques car il réduit l'abondance du microbiome, elles sont fragilisées
et plus sensibles à des bactéries pathogènes (Foucart 2018).
Les microbes symbiontes des insectes intéressent de plus en plus les
chercheurs. On connaissait déjà leur rôle dans la nutrition,
la digestion et la défense de l'hôte. Ils sont aussi sensibles
aux variations de température de leurs hôtes en fonction de l'environnement.
Ils aident la synthèse de cuticule et la sclérotisation qui permettent
la résistance à la dessication (voir de Souza et al 2011).
Ils peuvent aussi détoxifier des métaux lourds.
La deltaméthrine (insecticide
pyréthrinoïde, neurotoxique, très utilisé contre les
moustiques) réduit significativent la durée de vie des abeilles,
et aussi diminue la diversité du microbiome intestinal. Certaines bactéries
accroissent la résistance des abeilles à la deltaméthrine
en augmentant la production de défensine et de Toll (Dong et al 2022)
Chez les termites mangeurs de bois il y a évidemment un important microbiome. Celui-ci peut évoluer lors de migrations comme c'est le cas pour le termite Reticulitermes flavipes originaire de Louisianne et introduit en France au cours du 18ème siècle (Dedeine et al 2017). La socialité aurait été favorisée chez les insectes qui vivent dans le bois plus ou moins pourri, depuis les soins parentaux jusqu'à l'eusocialité des termites. Il est facile de transmettre les microbiotes aux descendants. Les interrelations comme les léchages interindividuels et le nettoyage du nid ont permis la lutte contre les microbes invasifs (Dillard et Benbow 2020).
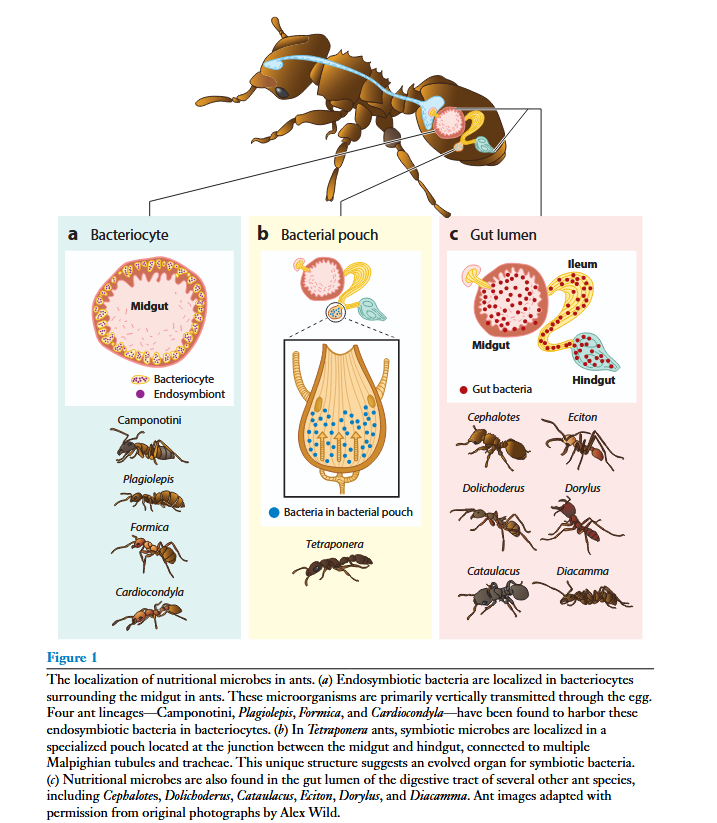
Les fourmis ne sont pas en reste puisque l'on y trouve de nombreuse bactéries.
Une revue faite sur 4 genres (Camponotus avec Blochmannia, Plagiolepis, Formica et Cardiocondyla) montre que le seul point commun est la capacité des bactéries à synthétiser la tyrosine. Cela signifie qu'il y a eu des évolutions parallèles indépendantes dans l'adoption des symbiontes par les fourmis (Jackson et al 2022).
- Chez Camponotus les bactéries symbiotes Blochmania ont été découvertes depuis plus d'un siècle mais leur rôle vient seulement d'être compris. En effet il est impossible de les élever comme des bactéries "normales". Chez C. fellah, on les trouve dans le tube digestif dans des cellules spécialisées appelées bactériocytes. Danival Souza à Tours pendant sa thèse a montré qu'elles facilitent la croissance de la colonie et les défenses immunitaires (Souza et al 2006, Souza et al 2009). Ces bactéries sont transmises par la reine depuis ses ovaires vers les oocytes (Kupper et al 2016). Si on traite les fourmis avec un antibiotique, elles vont réagir comme en situation de stress, produire plus d'hydrocarbures cuticulaires et devenir plus mélanisées (Souza et al 2011).
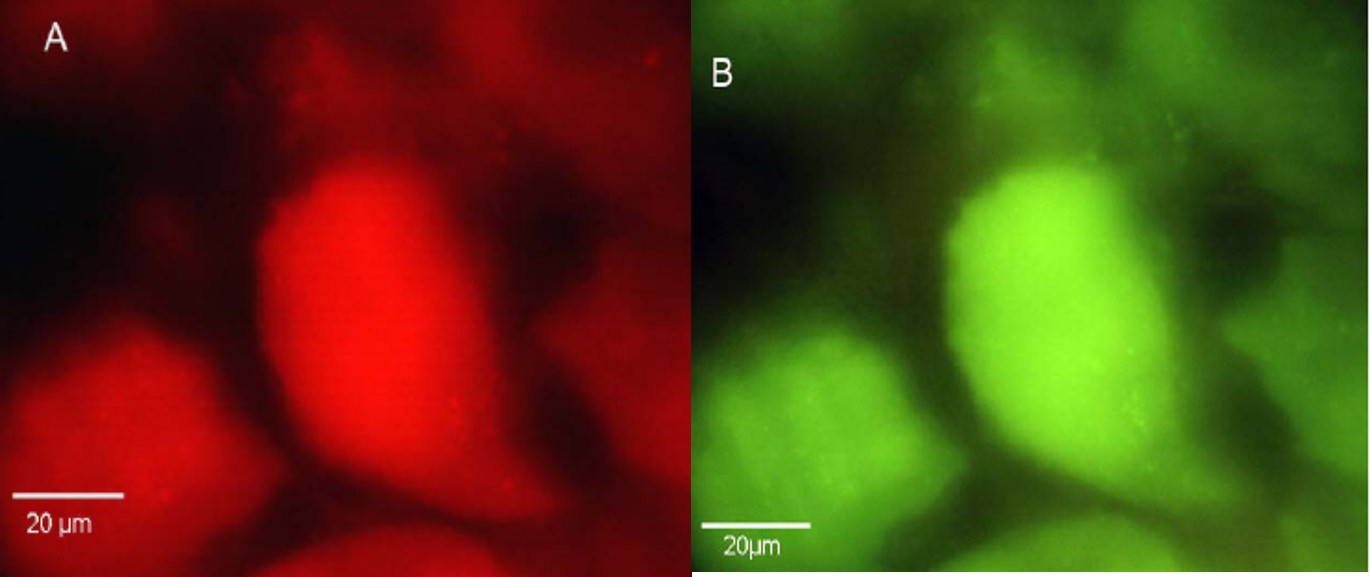
Les bactéries endosymbiotiques
Blochmannia ont évolué chez les fourmis Camponotus
mais se
retrouvent aussi chez Polyrachis et Colobopsis (Rafiqi et
al 2020, voir Nick 2020) :
"Les
scientifiques estiment que l’interdépendance entre la bactérie
Blochmannia et les fourmis Camponotini remonte à il y a 51 millions d’années.
Les chercheurs ont découvert que la bactérie entoure complètement
la lignée germinale. Le développement de l’embryon est parfaitement
organisé. Ainsi, les biologistes ont examiné de près les
gènes qui régulent la lignée germinale et le contenu génétique
de ces fourmis. L’équipe de scientifiques a constaté des
faits exceptionnels. Normalement, chez les insectes, les gènes de la
lignée germinale sont localisés à un seul endroit dans
l’ovule. Pourtant, dans le cas étudié, ils sont situés
sur quatre endroits différents. « Personne n’a jamais rien
vu de tel chez un autre insecte », a déclaré Arjuna Rajakumar,
coauteur de l’article. Par ailleurs, d’après ce qu’a
expliqué Matteen Rafiqi, biologiste à l’Université
Bezmialem Vakif à Istanbul, les gènes qui définissent la
structure du corps apparaissent normalement à un stade tardif du développement
de l’embryon. Or, chez le Camponotini, sont apparus très tôt
et se situent aux quatre mêmes endroits que les gènes de la lignée
germinale. Les scientifiques ont dû travailler avec plus de 30 espèces
de fourmis étroitement apparentées. Toutefois, ils ont réussi
à reconstituer les étapes de fusion. Ils ont retracé le
processus depuis l’époque où les gènes de la lignée
germinale étaient localisés dans un seul endroit, jusqu’à
ce qu’ils se retrouvent, avec les gènes Hox, dans quatre zones
différentes.
« Ces connaissances peuvent conduire à une meilleure compréhension
de l’origine des organismes complexes », a souligné Ehab
Abouheif, biologiste à l’Université McGill et auteur principal
de l’article.
« Ces étapes d’unification pour former une relation endosymbiotique
obligatoire nous aideront à comprendre d’autres fusions majeures
qui ont donné naissance à des formes de vie complexes, comme lorsque
des organismes unicellulaires se sont réunis pour former des organismes
multicellulaires », a-t-il confié."
Au cours de cette coévolution la fourmi (Camponotus) a même appris à contrôler les bactéries par autophagie (Gonçalves et al 2020).
On sait que les fourmis comme Camponotus floridanus utilisent l'acide formique comme arme défensive mais c'est aussi un antimicrobien. Elles s’arrosent d’acide formique juste après le repas. Elles en récupèrent en se toilettant et il va dans le jabot où il entretient le milieu acide favorable aux bactéries symbiontes. Durant les heures qui suivent, les fourmis se toilettent plus fréquemment au niveau de l’acidopore et le pH dans la lumière de leur intestin moyen augmente. Alimentées par un sirop contaminé par Serratia marcescens (bactérie pathogène), les ouvrières à l’acidopore bouché encourent un risque de mort accru, de même que celles à qui elles ont donné du régurgitat. Cela semble valable aussi pour d'autres Formicines (Camponotus maculatus, Lasius fuliginosus, Formica cinerea, F. cunicularia, F. pratensis et F. rufibarbis). Voir Pharmacon d'Alain Fraval (Tragust et al 2020).
Voir aussi (Demeure 2020)
:
"Chez les humains, l’estomac produit directement l’acide
gastrique, celui-ci permettant de réduire la taille des portions alimentaires
en dénaturant les protéines. Également, l’acide gastrique
tue la plupart des bactéries indésirables. Il s’agit d’une
solution d’acide chlorhydrique dont la concentration et le pH varient
selon la distance d’un repas. Chez les fourmis (Formicidae), il existe
un équivalent. Ces insectes produisent un poison à l’aide
de glandes spécifiques situées au niveau de l’abdomen.
Selon une étude publiée dans eLife le 3 novembre 2020, les fourmis
aspirent leur propre poison. Les chercheurs de l’Université de
Bayreuth (Allemagne) sont formels : cette technique permet aux fourmis de procéder
à une acidification de leur estomac. Le principal auteur de l’étude,
Simon Tragust, explique que ceci permet de limiter la prolifération des
microbes pathogènes avalés par accident. De plus, les fourmis
procéderaient à ce nettoyage après chaque repas. «Après
avoir accédé à l’acide, les fourmis voient leurs
chances de survie considérablement augmenter après avoir mangé
des aliments enrichis en bactéries pathogènes», a déclaré
Simon Tragust dans un communiqué."
- Il y a aussi des bactéries symbiotiques dans le tube digestif des Dolichoderus du Pérou. Ces fourmis sont herbivores, mangent du miellat de pucerons et du nectar extrafloral pauvres en azote. Les bactéries aident au recyclage de l'azote et la synthèse de vitamines et amino-acides. Au contraire chez Harpegnathos saltator carnivore, les gènes ne sont plus présents au profit de ceux qui permettent la transformation d'histidine et arginine en glutamine. Les bactéries sont des Bartollenaceae que l'on retrouve aussi chez l'abeille et qui provoquent des infections chroniques ou pathogènes chez les mammifères (Bisch et al 2018).
- Chez les fourmis champignonnistes on a découvert ces dernières années que la symbiose entre la fourmi et champignon cultivé est bien plus complexe que ce que l’on pensait. Des champignons parasites très virulents du genre Escovopsis peuvent se développer dans la culture et tuer la colonie rapidement. La réponse des fourmis a été de domestiquer des actinobactéries du genre Pseudonocardia qui se trouvent sur le corps de la fourmi et secrètent des antibiotiques (tâches blanches sur les photos ci-dessous). Un gros article de l'équipe de Cameron Curie a montré l'évolution des champignonnistes qui ont domestiqué ces bactéries pour se protéger des Escovopsis qui attaquent les champignons. L'un des antibiotiques a été identifié, il s'appelle dentigérumycine (Oh et al 2009); mais il ne semble pas qu'il soit intéressant pour l'homme. Ces bactéries secrètent d'autres antibiotiques comme la candicidine (connue depuis 1953) qui sont actifs aussi contre des Streptomyces (Haeder et al 2009, Dangelo et al 2016). Ces bactéries sont aussi emportées par la jeune reine fondatrice. Le système est encore bien plus complexe avec des levures qui mangent les bonnes bactéries ; et d’autres micro-organismes en cours de découverte. On vient par exemple de trouver des bactéries fixatrices d’azote comme celles qu’on trouve dans les racines de légumineuses. Il existe un véritable « microbiome » bactérien dans la meule à champignon où ce sont des bactéries qui digèrent les parois cellulaires des plantes (Suen et al. 2010). Les fourmis champignonnistes ont aussi une flore bactérienne importante dans leur estomac, mais elle est simple. Acromyrmex possède principalement seulement 4 taxa bactériens (Wolbachia, Rhizobiales et 2 Entoplasmatales). Les Rhizobiales sont uniquement extracellulaires dans l’intestin où elles forment des biofilms (ce qui les protège des antibiotiques comme la tétracycline) et possèdent des protéines permettant la fixation d’azote (Sapountzis et al. 2015).
 Actinobactéries
:
Actinobactéries
: 

Nathaniel Herzberg du Monde attire notre attention sur une publication récente "Contre l’antibiorésistance, les fourmis donnent l’exemple" : les fourmis Acromymex ont adopté des bactéries Pseudonocardia pour lutter contre d’autres bactéries, les Streptomyces, qui tuent le champignon symbiotique. (voir Pathak et al 2019).
Chez les Acromyrmex echiniator du Panama il y a de nombreuses bactéries dans le tube digestif (des Wolbachia, des entomoplasmales EntAcro, des rhizobiales). Le traitement avec de la tétracycline perturbe la production de deux acides gras (4-oxo-octanoic acide et 4-oxo-decanoic acide) de la glande métapleurale et modifie un peu le profil des hydrocarbures cuticulaires (pour le C36 et le C40). Cela entraîne des modifications dans la reconnaissance coloniale. Les auteurs ont même essayé de faire un traitement avec des goutelettes fécales de fourmis normales mais c'est seulement partiellement efficace ! (Teseo et al 2019);
Les Pseudonocardia sont des Actinobactéries symbiontes des fourmis champigonnistes qu'elles protègent avec des antifongiques et antibiotiques divers. Il y a des échanges de bactéries à tous les niveaux entre fourmis et surtout aussi avec des bactéries de l'environnement avec des transferts horizontaux. De quoi vous faire tourner la tête. On connait déjà près d'une vingtaine d'antibiotiques et antifongiques de Pseudonocardia (avec de jolis noms comme gérumycine, selvamycine..) et on espère en découvrir encore beaucoup d'autres (Goldstein et al 2020).
Trachymyrmex
avec Pseudonocardia
: 
Les types d'échanges
: 
- Chez les fourmis rousses en dôme de nos forêts, la chaleur du dôme serait fortement influencée par les bactéries qui se nourrissent de miellat et de résine (Jílková et al 2018).
- Les fourmis
Azteca qui vivent en symbiose avec les Cecropia (parasoliers)
logent dans les entre-nœuds de la plante. Les chambres ont des fonctions
déterminées : pouponnière, réserve de nourriture,
salle de repos, décharge. Chaque chambre de la domatie a un microbiome
particulier et différent de celui du milieu environnant. Les Azteca
maintiennent une propreté excellente, surtout autour du couvain. En revanche,
elles ne s’occupent pas des agents de maladies pouvant affecter le parasolier
; de ce point de vue, elles ne le protègent pas.
Les microbiomes varient selon l’endroit mais partout, la propreté
est bien supérieure à ce qu’on trouve dans nos maisons et
appartements.. Voir Lucas et al 2019 et Une
leçon de propreté.
- On vient aussi de découvrir que les fourmis influencent la flore bactérienne sur les feuilles de l'acacia. En absence de la fourmi Pseudomyrmex hindsii les feuilles sont plus attaquées par des pathogènes et présentent donc plus de dommages comme des trous ou des zones sombres. C'est aussi le cas si l'arbre est colonisé par une fourmi parasite Pseudomyrmex gracilis (on parle de parasite car la fourmi ne défend pas l'arbre contre les prédateurs). Ce fait est lié à des modifications de la flore bactérienne des feuilles. On a donc un effet indirect (González-Teuber et al 2014).
- M. Cooling et B. Hoffmann en Australie ont observé que entre 2003 et 2014 la fourmi folle jaune pouvait disparaître spontanément dans divers endroits (Cooling and Hoffmann 2015). Cette disparition pourrait être due à des virus, et d'autres bactéries pathogènes comme Rhabdochlamydia, Serratia et Cardinium (Cooling et al 2016).
- En 2017 on vient d'identifier toute une série d'antibiotiques efficaces contre deux bactéries résistantes aux antibiotiques. Ce sont des polypeptides aromatiques appelés fasamycins et formicamycins issus de bactéries filamenteuses actinomycètes. Ces bactéries ont été trouvées dans des nids de fourmis Tetraponea penzigi dans des domaties sur des acacias au Kénya (Qin et al 2017). On en trouve aussi chez des guêpes.
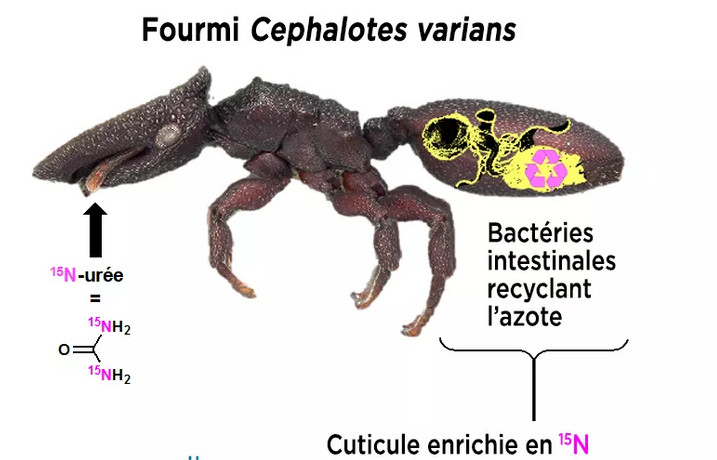
- Chez Cephalotes, fourmi tortue arboricole se nourrissant de nectar, de pollen, de champignons, régimes à faible teneur en azote nécessitant l'action de bactéries, le transfert des bactéries des fourmis les plus âgées aux plus jeunes se fait par secrétions anales ("transplantations fécales" selon Charpentier 2018, coprophagie selon Lanan et al).
Le
rôle important des bactéries chez les fourmis tortues Cephalotes
varians.
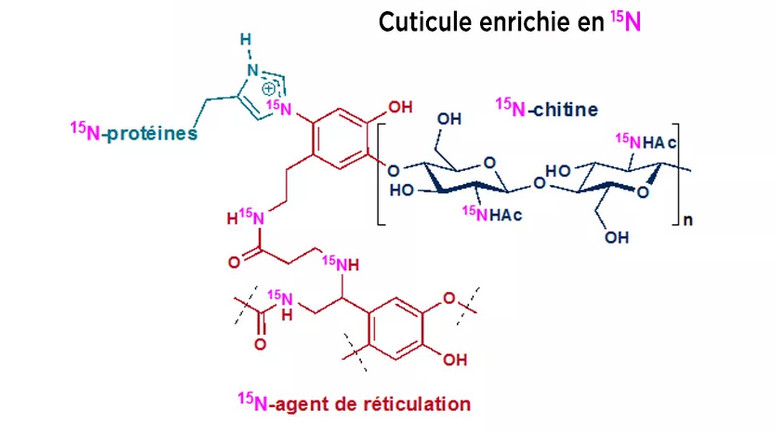
"En utilisant de l'urée marquée avec son isotope azote
15N qu'ils ont pu tracer par spectrométrie de masse et spectroscopie
RMN, les équipes ont montré que tous les composants cuticulaires
(protéines, chitine et agents de réticulation) étaient
enrichis en 15N, mettant en évidence l'apport en azote dans la cuticule
par bactéries symbiotiques qui réalisent la transformation de
l'urée. Ces résultats montrent que acides aminés produits
par les bactéries sont utilisés dans les différentes voies
de biosynthèse nécessaires à la formation de la cuticule.
Résultats qu'il faut maintenant tenter de relier aux propriétés
mécaniques de ces carapaces." (Article de Duplais et al 2021,
voir Adrien 2021).
- Les larves de Liometopum (escamole du Mexique) sont très recherchées pour leur goût (caviar des mexicains). Elles ont des bactéries symbiotes du type Firmicutes fixatrices d'azote, ce qui explique leur haute teneur en protéines, alors que les adultes ont surtout des Protéobactéries. Les bactéries des larves sont contenues dans des bactériocytes, elles servent à la synthèse des vitamines et des acides aminés (González-Escobar 2018). Voir Fourmis comestibles.
- Les Allomerus ont aussi dans leurs nids des actinobactéries (6 Streptomyces et 1 Amycolatopsis) qui les protègent contre les champignons parasites (Seipke et al 2012).
- Il est même possible que la production d'alcaloïdes soit liée à la présence de bactéries comme cela vient d'être montré chez les Aphaenogaster. A. senilis a de grandes quantités d'alcaloïdes dans sa glande à poison alors que A. iberica n'en a pas du tout. Si on traite les A. senilis avec un antibiotique, la production d'alcaloïdes chute considérablement (94%). Le traitement induit aussi un état de stress qui se traduit par une augmentation d'hydrocarbures cuticulaires. Le traitement n'a aucun effet sur A. iberica (Lenoir et Devers 2018). Cela vient d'être confirmé avec des bactéries du genre Serratia qui ont été trouvés chez les attines Atta et Acromyrmex, qui produisent des pyrazines (phéromones de piste de la glande à poison), à partir de thréonine (Silva-Junior et al. 2018). On vient aussi d'en trouver chez Myrmica scabrinodis (Scarparo et al 2020).
- Les fourmis Cataglyphis du désert de Tunisie ont des bactéries Weissella dans leur tube digestif que l'on trouve aussi dans les crottes de chameau. Ces dernières ont des propriétés anti-cholestérol et antibiotiques. On pourrait en faire peut-être des probiotiques (Fhoula et al 2018).
- Transferts de gènes entre fourmis et bactéries. Oui, la bactérie Streptomyces clavuligerus qui fabrique des peptides anti-microbiens aurait récupéré les gènes permettant leur synthèse à partir d'hyménoptères dont de nombreuses espèces de fourmis (Ayala-Runao et al 2019). Mais on n'a pas trouvé de transferts horizontaux entre la fourmi Formica sanguinea et ses esclaves Serviformica pour les symbiontes Wolbachia et Spiroplama ou Entomoplasma (Haapaniemi and Pamilo 2015).
- Le lait des fourmis. Une belle découverte de l'équipe de Kronauer : les pupes d'Oocera biroi secrètent lors de la mue imaginale un fluide riche en nutriments, hormones et substances neuroactives. Cette secrétion est rapidement consommée par les jeunes larves. Elle est cruciale pour la survie des pupes, si elle n'est pas consommée, des infections fongiques tuent la pupe. Cela peut être comparé au lait des mammifères (Snir et al 2022, voir commentaires de d'Ettorre et Tsuji 2022 et Herzberg 2022).
- La cuticule des fourmis héberge aussi bien sûr de nombreuses bactéries comme l'a montré Caroline Birer dans sa thèse (Birer 2017, Birer et al 2017, Birer et al 2020). Chez les espèces Atta cephalotes et Pseudomyrmex penetrator les résultats du métabarcoding ADN révèlent des différences inter- et intraspécifiques dans la composition des communautés bactériennes cuticulaires. Elle identifié un dipeptide cyclique (Cyclo(LPro-LPhe)) antimicrobien qui a été isolé à partir d’une souche proche de Streptomyces. La composition du microbiote bactérien cuticulaire des espèces Camponotus femoratus et Crematogaster levior dans les jardins de fourmis a été anlysée. Les résultats soulignent l’acquisition d’une partie du microbiote à partir de l’environnement. En parallèle l’analyse métabolomique des cuticules montre à contrario une plus grande spécificité liée à l’espèce de fourmi.
Chez les autres Arthropodes : on trouve des bactéries un peu partout quand on les cherche.
- Le microbiome des coléoptères myrmécophiles des nids de Formica polyctena semble très varié. On y trouve comme chez les fourmis des Wolbacchia et des Rickettsia (Kaczmarczyk-Ziemba et al 2020).
- De nombreuses punaises sont porteuses de Wolbachia. Suite à un traitement antibiotique, les punaises produisent moins de substances défensives et de phéromones d’alarme (Becerra et al. 2015).
- L'odeur coloniale des termites est influencée par les bactéries du tube digestif (Matsuura 2001, Minkley et al 2006).
- Chez les isopodes les Wolbachia protègent contre des infections de microbes pathogènes intracellulaire (Braquart-Varnier et al. 2015). Chez Armadillidium les bactéries symbiotiques Wolbachia ont un rôle dans la mise en place de l’odeur cuticulaire qui permet le choix d’orientation sexuelle, les mâles savent différencier des femelles infectées ou non (Richard 2017.
- Les abeilles
ont aussi évidemment une flore intestinale importante avec des microbes
capables de digérer le pollen et le nectar. Les abeilles avec une flore
variée résistent mieux à certains parasites (Enders 2015
p.214). Chez
l'abeille domestique on trouve 8 phylotypes de bactéries intestinales.
En Chine, on a traité des larves d'ouvrières en
milieu stérile de manière à les rendre
déficientes en bactéries du microbiote intestinal.
Les abeilles sans bactéries font 28 fois moins d'apidacéine, une
protéine antimicrobienne, que celle à qui on avait ensuite inoculé
des bactéries de tube digestif. Elles sont moins résistantes à
une infection par Escherichia coli (Nature 2017; Kwong et al 2017).
Chez les abeilles sans microbiote intestinal l'expression de certains gènes
du système immunitaire est affaiblie (apidacéine, abacéine,
hyménoptacéine et JNK, mais pas des défensines) et aussi
pour la vitellogénine. Par ailleurs, les bactéries intestinales
inhibent la prolifération du Nosema ceranae (parasite microsporide)
(Wu et al 2020).
Le microbiote participe à la mise en place de l'odeur coloniale avec
les hydrocarbures et
donc à l'identité coloniale. Chaque colonie possède un
microbiome spécifique qui influe sur les hydrocarbures. Pour le moment
on ne sait pas comment cela se réalise, peut-être en modifiant
l’expression des enzymes utilisées dans la synthèse des
hydrocarbures, ou en fournissant différents composés aux œnocytes
où se fait la synthèse (Vernier et al 2020, voir Rohrbacher 2020).
Chez certains coléoptères cette perturbation du microbiome perturbe
la synthèse d'acides aminés comme la tyrosine qui intervient dans
la synthèse des éléments de la cuticule (mélanisation
et sclérotisation) (Kiefer et al 2021) et donc influe sur le profil cuticulaire.
- Chez
les bourdons. En situation d'apprentisssage, chez Bombus
impatiens, il n'y a pas de différence entre les bourdons en élevage
avec ou sans microbiote intestinal (Leger et al 2020).
Lars Chittka, le grand spécialiste anglais des bourdons, travaille avec
les chinois. Ils ont exploré le microbiome intestinal de bourdons Bombus
terrestris et trouvé une corrélation positive entre la mémoire
d'une discrimination visuelle et la quantité de lactobacilles Firm-5,
mais pas avec les autres lactobacilles. Si on supplémente des bourdons
avec ce bacille on trouve plus de LPA (un glycérophospholipide, composant
des membranes biologiques) dans l'hémolymphe. De même la
supplémentation orale de LPA accroit la mémoire (Li et al 2021).
Signatures
génomiques de bourdons
Une étude du génome de mâles de Bombus terrestris
en Gde-Bretagne. Les auteurs ont observé une très forte variabilité
du génome concernant la neurobiologie et le développement des
ailes. Ils ont aussi trouvé un transfert horizontal de gènes avec
des bactéries Wolbachia (Colgan et al 2022).
- Chez les drosophiles les bactéries commensales (surtout Wolbachia et Lactobacillus) jouent un rôle dans le choix de l'espèce du partenaire sexuel par l'intermédiaire des hydrocarbures cuticulaires, ce qui peut jouer un rôle dans la spéciation (Sharon et al. 2010).
- Chez 25 espèces de guêpes Philanthus prédatrices d'abeilles (beewolf wasps) on trouve 49 antibiotiques contre les Streptomyces avec des variations entre espèce
Chez les vertébrés
Le rôle du microbionte devient fondamental dans la biologie des organismes comme le signale Science & Vie de janvier 2021 (Sciamma 2021)
Les bactéries peuvent participer à l’émission de signaux de reconnaissance comme chez les hyènes où l’odeur individuelle est plus variable entre groupes qu’à l’intérieur du groupe (Theis et al. 2012; Theis et al. 2013).
Voir
- Charpentier,
J. (2018) Les fourmis Céphalotes et leur symbiose bactérienne
de 46 millions d’années. actualite.housseniawriting.com, p. https://actualite.housseniawriting.com/science/biologie/fourmis/2018/03/07/les-fourmis-cephalotes-et-leur-symbiose-bacterienne-de-46-millions-dannees/26053/.
Pdf
- de Souza, D. J., S. Devers and A. Lenoir (2011). Blochmannia endosymbionts
and their host, the ant Camponotus fellah: Cuticular hydrocarbons and melanization.
Comptes Rendus Biologie 334: 737-741. Pdf
- Fraval, A. (2017)
Petites chimistes. Inra.fr/opie-insectes 2017. http://www7.inra.fr/opie-insectes/epingle17.htm
Voir
-
Herzberg, N. (2018). Les fourmis, premières agricultrices bio. Le Monde
7 octobre 2018. Voir
texte
- Lenoir, A. and S. Devers (2018). Alkaloid secretion
inhibited by antibiotics in Aphaenogaster ants. Comptes Rendus Biologies 341:
358-361. https://doi.org/10.1016/j.crvi.2018.06.004Pdf
- de Souza, D.J., A. Bézier, D. Depoix, J.-M. Drezen and A. Lenoir (2009).
Blochmannia endosymbionts improve colony growth and immune defence
in the ant Camponotus fellah. BMC Microbiology 9(1): 29. Pdf
- de Souza, D. J., D. Depoix, J. Lesobre and A. Lenoir (2006). Camponotus
fellah (Formicidae : Formicinae) et son endosymbionte, Candidatus Blochmannia.
Union internationale pour l’étude des insectes sociaux –
section française. Colloque annuel. Avignon – 24-27 avril 2006:
62. Pdf
- Adrien (2021) Comment se forger une carapace ? techno-science.net
16 février 2021. https://www.techno-science.net/actualite/comment-se-forger-carapace-N20574.html
- Artavia-León,
A., M. Pacheco-Leiva, C. Moya-Román, N. Rodríguez-Hernández
and A. A. Pinto-Tomás (2019). Ant microbial symbionts are a new model
for drug discovery. Drug Discovery Today: Disease Models. https://doi.org/10.1016/j.ddmod.2019.08.011
- Ayala-Ruano, S., D. Santander, E. Tejera, Y. P. Castillo and V. D. Armijos
Jaramillo (2019). A putative antimicrobial peptide from Hymenoptera in the megaplasmid
pSCL4 of Streptomyces clavuligerus ATCC 27064 reveals a singular case of horizontal
gene transfer with potential applications. Ecology and Evolution sous presse.
DOI: 10.1002/ece3.4924
- Barnéoud, L. and A. Bomboy (2018). L'Inde, fabrique d'antibiorésistance.
Le Monde Science & Médecine 12 décembre 2018. p. 1, 4-5.
- Barthélémy, P. (2016). Il n'y a pas d'argent qui ne soit pas
sale. Le Monde Science et Médecine 21 décembre 2016.
- Becerra, J., G. Venable and V. Saeidi (2015). Wolbachia-Free Heteropterans
Do Not Produce Defensive Chemicals or Alarm Pheromones. Journal of Chemical
Ecology 41(7): 593-601. 10.1007/s10886-015-0596-4
- Birer, C. (2017) Le microbiote bactérien cuticulaire des fourmis de
Guyane : pouvoir antibiotique et écologie des communautés. https://tel.archives-ouvertes.fr/tel-01537793
- Birer, C., N. Tysklind, L. Zinger and C. Duplais (2017). Comparative analysis
of DNA extraction methods to study the body surface microbiota of insects: A
case study with ant cuticular bacteria. Molecular Ecology Resources 17(6). 10.1111/1755-0998.12688
- Birer C., CS Moreau, N Tysklind, L Zinger, C Duplais (2020). Disentangling
the assembly mechanisms of ant cuticular bacterial communities of two Amazonian
ant species sharing a common arboreal nest. Molecular ecology 29 (7), 1372-1385
- Bisch, G., M. M. Neuvonen, N. Pierce, J. A. Russell, R. Koga, J. G. Sanders,
P. Lukasik and S. G. E. Andersson (2018). Genome Evolution of Bartonellaceae
Symbionts of Ants at the Opposite Ends of the Trophic Scale. Genome Biol Evol
10: 1687-1704.
- Braquart-Varnier, C., M. Altinli, R. Pigeault, F. Chevalier, P. Greve, D.
Bouchon and M. Sicard (2015). The mutualistic side of Wolbachia-Isopod interactions:
Wolbachia mediated protection against pathogenic intracellular bacteria. Frontiers
in Microbiology 6. 10.3389/fmicb.2015.01388
- Cabut, S. and N. Herzberg (2017). Le microbe, un indice au service de la police.
Le Monde 23 août 2017. p. 7.
-
Colgan, T. J., A. N. Arce, R. J. Gill, A. Ramos Rodrigues, A. Kanteh, E. J.
Duncan, L. Li, L. Chittka and Y. Wurm (2022). Genomic Signatures of Recent Adaptation
in a Wild Bumblebee. Molecular Biology and Evolution 39(2). doi: 10.1093/molbev/msab366.
- Cooling, M. and B. D. Hoffmann (2015). Here today, gone tomorrow: declines
and local extinctions of invasive ant populations in the absence of intervention.
Biological Invasions 17(12): 3351-3357. 10.1007/s10530-015-0963-7
- Cooling, M., M. A. M. Gruber, B. D. Hoffmann, A. Sébastien and P. J.
Lester (2016). A metatranscriptomic survey of the invasive yellow crazy ant,
Anoplolepis gracilipes, identifies several potential viral and bacterial pathogens
and mutualists. Insectes Sociaux: 1-11. 10.1007/s00040-016-0531-x
- d’Ettorre, P. and K. Tsuji (2022). A fluid role in ant society as adults
give larvae ‘milk’ from pupae. Nature 30 November 2022. doi: 10.1038/d41586-022-03722-4.
.
- Dangelo, R. A. C., D. J. De Souza, T. D. Mendes, J. d. C. Couceiro and
T. M. C. Della Lucia (2016). Actinomycetes inhibit filamentous fungi from the
cuticle of Acromyrmex leafcutter ants. J. Basic Microbiol. 56: 1-9.
- Dedeine, F., S. Bankhead-Dronnet, C. Lopez-Vaamonde and C. Bouget (2017).
Invasions biologiques : le rôle des microorganismes symbiotiques. Microscoop
HS, octobre 2017: 16-17.
- Demeure, Y. (2020) Les fourmis ont une surprenante manière de se
prémunir des infections ! sciencepost.fr,
22 novembre 2020.
- Dillard, J. and M. E. Benbow (2020). From Symbionts to Societies: How
Wood Resources Have Shaped Insect Sociality. Frontiers in Ecology and Evolution
8(173). doi: 10.3389/fevo.2020.00173.
-
Dong, Z.-X., Q.-H. Tang, W.-L. I. Li, Z.-W. Wang, X.-J. Li, C.-M. Fu, D. Li,
K. Qian, W.-L. I. Tian and J. Guo (2022). Honeybee (Apis mellifera) resistance
to deltamethrin exposure by Modulating the gut microbiota and improving immunity.
Environmental Pollution 314: 120340. doi: https://doi.org/10.1016/j.envpol.2022.120340.
- Duplais, C., V. Sarou-Kanian, D. Massiot, A. Hassan, B. Perrone, Y. Estevez,
J. T. Wertz, E. Martineau, J. Farjon, P. Giraudeau and C. S. Moreau (2021).
Gut bacteria are essential for normal cuticle development in herbivorous turtle
ants. Nature Communications 12(1): 676. doi: 10.1038/s41467-021-21065-y.
- Enders, G. (2015). Le charme discret de l'intestin, Actes Sud.
- Engl, T., J. Kroiss, M. Kai, T. Y. Nechitaylo, A. Svatoš and M. Kaltenpoth
(2018). Evolutionary stability of antibiotic protection in a defensive symbiosis.
Proceedings of the National Academy of Sciences. 10.1073/pnas.1719797115
- Fhoula, I., A. Rehaiem, A. Najjari, D. Usai, A. Boudabous, L. A. Sechi and
O. Hadda-Imene (2018). Functional Probiotic Assessment and In Vivo Cholesterol-Lowering
Efficacy of Weissella sp. Associated with Arid Lands Living-Hosts. BioMed Research
International 2018: 11. 10.1155/2018/1654151
- Foucart, S. (2018). Le glyphosate, autre cause possible du déclin des
abeilles domestiques. Le Monde 28 septembre 2018. p. 8.
- Goldstein, S. L. and J. L. Klassen (2020). Pseudonocardia Symbionts of Fungus-Growing
Ants and the Evolution of Defensive Secondary Metabolism. Frontiers in Microbiology
11: 621041-621041. doi: 10.3389/fmicb.2020.621041
- González-Teuber, M., M. Kaltenpoth and W. Boland (2014). Mutualistic
ants as an indirect defence against leaf pathogens. The New Phytologist 202(2):
640-650.
- González-Escobar, J. L., A. Grajales-Lagunes, A. Smolinski, A. Chagolla-López,
A. De Léon-Rodríguez and A. P. Barba De La Rosa (2018). Microbiota
of edible Liometopum apiculatum ant larvae reveals potential functions related
to their nutritional value. Food Research International in press. https://doi.org/10.1016/j.foodres.2018.04.04
- Gonçalves, W. G., K. M. Fernades, A. P. A. Silva, D. G. Gonçalves,
M. Fiaz and J. E. Serrao (2020). Ultrastructure of the Bacteriocytes in the
Midgut of the Carpenter ant Camponotus rufipes: Endosymbiont Control by Autophagy.
Microscopy and Microanalysis: 1-9. doi: doi:10.1017/S1431927620024484.
- Haapaniemi, K. and P. Pamilo (2015). Social parasitism and transfer of symbiotic
bacteria in ants (Hymenoptera: Formicidae). Myrmecological News 21: 49-57.
- Haeder, S., R. Wirth, H. Herz and D. Spiteller (2009). Candicidin-producing
Streptomyces support leaf-cutting ants to protect their fungus garden against
the pathogenic fungus Escovopsis. Proceedings of the National Academy of Sciences
106(12): 4742-4746. 10.1073/pnas.0812082106
- Herzberg, N. (2022) Les pupes de fourmis fabriquent un lait magique pour la
colonie. Le
Monde 4 décembre 2022. Avec interviews de Patrizia d'Ettorre
et Laurent Keller.
- Jackson,
R., D. Monnin, P. A. Patapiou, G. Golding, H. Helanterä, J. Oettler, J.
Heinze, Y. Wurm, C. K. Economou, M. Chapuisat and L. M. Henry (2022). Convergent
evolution of a labile nutritional symbiosis in ants. The Isme Journal. doi:
10.1038/s41396-022-01256-1
- Kiers, E. T. and S. A. West (2015). Evolving new organisms via symbiosis.
Science 348(6233): 392-394. 10.1126/science.aaa9605
- Jílková, V., T. Cajthaml and J. Frouz (2018). Relative importance
of honeydew and resin for the microbial activity in wood ant nest and forest
floor substrate – a laboratory study. Soil Biology and Biochemistry 117(Supplement
C): 1-4. https://doi.org/10.1016/j.soilbio.2017.11.002
- Kaczmarczyk-Ziemba, A., M. Zagaja, G. K. Wagner, E. Pietrykowska-Tudruj and
B. Staniec (2020). First Insight into Microbiome Profiles of Myrmecophilous
Beetles and Their Host, Red Wood Ant Formica polyctena (Hymenoptera: Formicidae)—A
Case Study. Insects 11(2): 134.
- Kiefer, J. S. T., S. Batsukh, E. Bauer, B. Hirota, B. Weiss, J. C. Wierz,
T. Fukatsu, M. Kaltenpoth and T. Engl (2021). Inhibition of a nutritional endosymbiont
by glyphosate abolishes mutualistic benefit on cuticle synthesis in Oryzaephilus
surinamensis. Communications Biology 4(1): 554. doi: 10.1038/s42003-021-02057-6
- Kupper, M., C. Stigloher, H. Feldhaar and R. Gross (2016). Distribution of
the obligate endosymbiont Blochmannia floridanus and expression analysis of
putative immune genes in ovaries of the carpenter ant Camponotus floridanus.
Arthropod Structure & Development. http://dx.doi.org/10.1016/j.asd.2016.09.004
- Kwong, W. K., A. L. Mancenido and N. A. Moran (2017). Immune system stimulation
by the native gut microbiota of honey bees. Royal Society Open Science 4(2).
10.1098/rsos.170003
- Lanan, M. C., P. A. P. Rodrigues, A. Agellon, P. Jansma and D. E. Wheeler
(2016). A bacterial filter protects and structures the gut microbiome of an
insect. The Isme Journal 10: 1866. 10.1038/ismej.2015.264. https://www.nature.com/articles/ismej2015264#supplementary-information.
Libre de droits
- Leger, L. and Q. S. McFrederick (2020). The Gut–Brain–Microbiome
Axis in Bumble Bees. Insects 11: 517.
- Lemoine, M., T. Engl and M. Kaltenpoth (2020). Microbial symbionts expanding
or constraining abiotic niche space in insects. Current Opinion in Insect Science.
10.1016/j.cois.2020.01.003
- Li, H., J. Sosa-Calvo, H. A. Horn, M. T. Pupo, J. Clardy, C. Rabeling,
T. R. Schultz and C. R. Currie (2018). Convergent evolution of complex structures
for ant–bacterial defensive symbiosis in fungus-farming ants. Proceedings
of the National Academy of Sciences. 10.1073/pnas.1809332115
- Li, L., C. Solvi, F. Zhang, Z. Qi, L. Chittka and W. Zhao (2021). Gut microbiome
drives individual memory variation in bumblebees. Nature Communications 12(1):
6588. doi: 10.1038/s41467-021-26833-4
- Lucas, J. M., A. A. Madden, C. A. Penick, M. J. Epps, P. R. Marting, J. L.
Stevens, D. J. Fergus, R. R. Dunn and E. K. Meineke (2019). Azteca ants maintain
unique microbiomes across functionally distinct nest chambers. Proceedings of
the Royal Society B: Biological Sciences 286(1908): 20191026. doi:10.1098/rspb.2019.1026
Libre
de droits
- Luk, F. L. (2018) Le
tour du monde des microbes. lejournal.cnrs.fr, 11 janvier 2018
- Matarrita-Carranza, B., R. D. Moreira-Soto, C. Murillo-Cruz, M. Mora,
C. R. Currie and A. A. Pinto-Tomas (2017). Evidence for Widespread Associations
between Neotropical Hymenopteran Insects and Actinobacteria. Frontiers in Microbiology
8(2016). 10.3389/fmicb.2017.02016. libre de droits
- Matsuura, K. (2001). Nestmate recognition mediated by intestinal bacteria
in a termite, Reticulitermes speratus. Oikos 92: 20-26.
- Minkley, N., A. Fujita, A. Brune and W. H. Kirchner (2006). Nest specificity
of the bacterial community in termite guts (Hodotermes mossambicus). Insectes
Sociaux 53: 339-344.
- Nature (2017). Microbiology: Gut bacteria boost bee immunity. Nature 542(7641):
274-274. 10.1038/542274a
- Nick (2020) On en sait plus sur l'origine des formes de vie complexes. fredzone.org.
10 septembre 2020. Lien
- Oh, D.-C., M. Poulsen, C. R. Currie and J. Clardy (2009). Dentigerumycin:
a bacterial mediator of an ant-fungus symbiosis. Nature Chemical Biology 5 (6):
391.
- Picq P. (2013) De Darwin à Lévy-Strauss. L’homme et la
diversité en danger , Odile Jacob
- Pathak, A., S. Kett and M. Marvasi (2019). Resisting Antimicrobial Resistance:
Lessons from Fungus Farming Ants. Trends in Ecology & Evolution. https://doi.org/10.1016/j.tree.2019.08.007
- Rafiqi, A. M., A. Rajakumar and E. Abouheif (2020). Origin and elaboration
of a major evolutionary transition in individuality. Nature 585: 1-6. doi: 10.1038/s41586-020-2653-6.
- Reeves, D. D., S. L. Price, M. O. Ramalho and C. S. Moreau (2020). The Diversity
and Distribution of Wolbachia, Rhizobiales, and Ophiocordyceps
Within the Widespread Neotropical Turtle Ant, Cephalotes atratus (Hymenoptera:
Formicidae). Neotropical Entomology. 10.1007/s13744-019-00735-z.
- Richard, F.-J. (2017). Symbiotic Bacteria Influence the Odor and Mating Preference
of Their Hosts. Frontiers in Ecology and Evolution 5(143). 10.3389/fevo.2017.00143.
Libre de droits
- Rohrbacher, F. (2020). Les abeilles domestiques s'identifient grâce
à leur microbiome. Le Monde Science et Médecine. 28 octobre 2020.
Pdf (voir texte)
- Sapountzis, P., M. Zhukova, L. H. Hansen, S. J. Sorensen, M. Schiott and J.
Boomsma (2015). Acromyrmex Leaf-Cutting Ants Have Simple Gut Microbiota with
Nitrogen-Fixing Potential Appl. Environ. Microbiol. 81: 5527-5537.
- Scarparo, G., P. Rugman-Jones, M. Gebiola and A. Di Giulio (2020). First screening
of bacterial communities of Microdon myrmicae and its ant host: do microbes
facilitate the invasion of ant colonies by social parasites? Basic and Applied
Ecology. doi: 10.1016/j.baae.2020.11.008.
- Sciama, Y. (2021). Bactéries. Elles font aussi ce que nous
sommes. Science & Vie. n°1240, janvier 2021. p.68-87.
- Sharon, G., D. Segal, J. M. Ringo, A. Hefetz, I. Zilber-Rosenberg and E. Rosenberg
(2010). Commensal bacteria play a role in mating preference of Drosophila melanogaster.
Proceeding of the National Academy of Science of USA 107: 20051-20056.
- Seipke, R. F., J. Barke, M. X. Ruiz-Gonzalez, J. Orivel, D. W. Yu and M. I.
Hutchings (2012). Fungus-growing Allomerus ants are associated with antibiotic-producing
actinobacteria. Antonie van Leeuwenhoek 101(2): 443-447. 10.1007/s10482-011-9621-y
- Silva-Junior, E. A., A. C. Ruzzini, C. R. Paludo, F. S. Nascimento, C. R.
Currie, J. Clardy and M. T. Pupo (2018). Pyrazines from bacteria and ants: convergent
chemistry within an ecological niche. Scientific Reports 8(1): 2595. 10.1038/s41598-018-20953-6
- Snir, O., H. Alwaseem, S. Heissel, A. Sharma, S. Valdés-Rodríguez,
T. S. Carroll, C. S. Jiang, J. Razzauti and D. J. C. Kronauer (2022). The pupal
moulting fluid has evolved social functions in ants. Nature. doi: 10.1038/s41586-022-05480-9.
- Suen, G., J. J. Scott, F. O. Aylward, S. M. Adams, S. G. Tringe, A. n. A.
Pinto-Tomás, C. E. Foster, M. Pauly, P. J. Weimer, K. W. Barry,
et al. (2010). An Insect Herbivore Microbiome with High Plant Biomass-Degrading
Capacity. PLoS Genet 6(9): e1001129.
- Teseo, S., J. S. van Zweden, L. Pontieri, P. W. Kooij, S. Sørensen,
T. Wenseleers, M. Poulsen, J. J. Boomsma and P. Sapountzis (2019). The scent
of symbiosis: gut bacteria may affect social interactions in leaf-cutting ants.
Anim Behav 150: 239-254. 10.1016/j.anbehav.2018.12.017
- Theis, K. R., T. M. Schmidt and K. E. Holekamp (2012). Evidence for a bacterial
mechanism for group-specific social odors among hyenas. Science Reports 2: 615.
- Theis, K. R., A. Venkataraman, J. A. Dycus, K. D. Koonter, E. N. Schmitt-Matzen,
A. P. Wagner, K. E. Holekamp and T. M. Schmidt (2013). Symbiotic bacteria appear
to mediate hyena social odors. Proceedings of the National Academy of Sciences.
10.1073/pnas.1306477110
- Tragust, S., C. Herrmann, J. Häfner, R.
Braasch, C. Tilgen, M. Hoock, M. A. Milidakis, R. Gross and H. Feldhaar (2020).
Formicine ants swallow their highly acidic poison for gut microbial selection
and control. eLife: 9:e60287. doi: 10.1101/2020.02.13.947432
- Vernier, C. L., I. M. Chin, B. Adu-Oppong, J. J. Krupp, J. Levine, G. Dantas
and Y. Ben-Shahar (2020). The gut microbiome defines social group membership
in honey bee colonies. Science Advances 6(42): eabd3431. doi: 10.1126/sciadv.abd3431.
- Wu, Y., Y. Zheng, Y. Chen, G. Chen, H. Zheng and F. Hu (2020). Apis cerana
gut microbiota contribute to host health though stimulating host immune system
and strengthening host resistance to Nosema ceranae. Royal Society Open Science
7(5): 192100. doi: doi:10.1098/rsos.192100.
- Zilberg-Rosenberg, I. and E. Rosenberg (2008). Role of microorganisms in the
evolution of animals and plants: the hologenome theory of evolution. FEMS Microbiology
Reviews 32: 723-735.
- Zhu, Y.-G., M. Gillings, P. Simonet, D. Stekel, S. Banwart and J. Penuelas
(2017). Microbial mass movements. Science 357(6356): 1099-1100. 10.1126/science.aao3007