
« Des fourmis adeptes de la mondialisation » selon Luc Passera
Alain Lenoir - Mis à jour le 28-Jui-2025
Espèces : Fourmi d'Argentine (Linepithema humile) - Fourmi de feu (Solenopsis invicta) - Fourmi folle jaune (Anoplolepis gracilipes) - Fourmi à grosse tête (Pheidole megacephala) - Fourmi électrique (Wasmannia auropunctata) - Fourmi fantôme (Tapinoma melanocephalum) - Fourmi des pharaons (Monomorium pharaonis) - Fourmi folle noire (Paratrechina longicornis) - Fourmi folle rousse (Nylanderia fulva) - Lasius neglectus - Fourmi rouge (Myrmica rubra) - D'autres espèces -
Thèmes : Caractéristiques des invasives - Pourquoi des fourmis deviennent-elles invasives ? - Compétition entre invasives - Invasives et lutte biologique - Des invasives qui disparaissent - Le coût des espèces de fourmis invasives - Lutte contre les fourmis invasives -
Invasives ou envahissantes ? Selon Alice Fournier (la biodiversité en péril, La Recherche n° spécial déc19-févr20) on peut séparer les deux, mais je parlerai le plus souvent d'invasives ...
Dans
Les
fourmis, de Sabine
Boccador, Fleurus 2021 : 
La mondialisation a des conséquences importantes
pour l'homme. De nombreuses maladies, transmises d'humain à humain (virus,
hépatite, rougeole..) ou par des insectes comme les moustiques (dengue,
chikungunya, paludisme..) se propagent très rapidement dans le monde
entier. Frédéric Joignot (2018a) estime que cette mondialisation
a commencé en 1493 avec l'arrivée de Christophe Colomb aux Caraïbes,
les espagnols apportant variole, grippe et rougeole. Il écrit que "La
mondialisation est aussi biologique". Il écrit aussi que
le plastique est devenu "une nouvelle espèce invasive",
en particulier dans les océans (2018b). En Nouvelle-Zélande les
colons ont introduits de nombreuses espèces comme des bourdons (Goulson)
ou la guêpe Vespula vulgaris qui attaque les fourmis Prolasius
advenus (Sciences et Avenir mai 2011).
Voir plus sur les envahiseurs dans le livre de Vincent Albouy "Etonnants
envahisseurs". Nous sommes de plus en plus confrontés à
des envahisseurs. C'est ainsi que chez nous, dans le Béarn, il y a un
rossignol du Japon échappé d'une volière qui s'est implanté
et vit très bien (voir photo plus
loin). Les
invasions biologiques
ne datent pas d'aujourd'hui
! On découvre par exemple que notre moineau domestique a envahi les Amériques
à la fin du 19ème siècle avec les colons européens
après avoir été considéré comme très
nuisible en Europe dans les annés 1830. Il est d'ailleurs en dégringolade
dans les villes, par exemple à Paris en 13 ans (de 2003 à 2016)
sa population a chuté de 75% (La Hulotte n°112, nov 2021).
Une
nouvelle synthèse sur les invasions biologiques
dans le monde (Angulo et al 2021). Analyse sur 243 pays pour toutes les espèces
invasives (animaux et plantes). Le record est pour les USA (hors outre-mer)
avec 523 espèces. La Polynésie, La Réunion et la Nouvelle-Calédonie
sont entre 170 et 190 espèces. Tapinoma melanocephalum (fourmi
fantôme), arrive à 40% des pays. Cette espèce se disperse
comme IP = Ignorant possessions = passagers clandestins. Rappelons le travail
d'Arnan et al (2021) qui montre que les fourmis invasives occupent en Europe
des niches climatiques vides.
Les espèces invasives de fourmis mettent en péril les écosystèmes du monde entier. De Rebecca Dzombak (National Geographic, 25 janvier 2023)
La
notion de supercolonie.
Une revue par H. Helanterä.
La définition de supercolonialité n'est pas claire.
- Elles se reproduisent par fission des colonies ("budding") ce qui
conduit à former de la polydomie et des déplacements d'ouvrières
entre nids.
- Les accouplements sont intranidaux ou localisés près des nids,
ce qui augmente la polygynie.
- Elles sont écologiquement dominantes
- Pourtant, souvent il n'y a qu'une faible parentèle entre les
individus, ce qui laisse la place à de la compétition. On a des
exemples de larves de Formica cannibales envers des oeufs. Cela pourrait
être une explication à la rareté des espèces supercoloniales,
mais en même temps les avantages expliquent le maintien de ce type de
colonialité.
Néanmoins elles ne sont pas toutes invasives et toutes les fourmis invasives
ne sont pas supercoloniales.
L'auteur fait un tableau récapitulatif des espèces supercoloniales.
La fourmi d'Argentine forme les supercolonies les plus grandes connues sur plusieurs
continents. Pour les Tapinoma il y a T. sessile et T.
melanocephalum, mais pas encore T. magnum. En Europe il y a par
exemple Lasius neglectus, Monomorium pharaonis.
Le
coût des espèces invasives
Gros travail sur le coût des invasions biologiques
animé par Franck
Courchamp publié dans Nature (Diagne et al 2021) sur la
littérature en anglais et Science of the Total Environment sur la littérature
dans des langues autres que l'anglais (Angulo
et al 2021).
Sur le premier article, selon Pham (2021) : "Une étude
chiffre les dégâts causés par les espèces envahissantes
à près de 1300 milliards de dollars pour ces 40 dernières
années. Introduites volontairement ou non par l’homme dans un nouveau
milieu, les espèces envahissantes sont des espèces exotiques qui
deviennent nuisibles dans leur nouvel habitat, explique au HuffPost Franck Courchamp,
co-auteur de l’étude et directeur de recherche au CNRS. Et
de plus en plus d’invasions d’espèces surviennent, déclare
le chercheur. En cause d’abord, la croissance des échanges de marchandises
qui accompagnent l’accélération de la mondialisation. “Une
espèce invasive de fourmis peut voyager avec des plantes ornementales,
des larves de poissons peuvent être introduites dans les eaux de ballast
des navires”, avance Franck Courchamp. Le réchauffement climatique
joue également un rôle, en facilitant la survie d’espèces
importées de régions plus chaudes du globe." "Aux Etats-Unis,
rien que les attaques des fourmis de feu entraînent une centaine de milliers
d’hospitalisations et 100 décès par an, pour un coût
annuel de 6 milliards de dollars."
En ce qui concerne la littérature dans 15 langues autres que l'anglais,
c'est très impressionnant, il y encore plus de données (2550 vs
2306 pour les coûts), cela ajoute 249 espèces invasives et 15 pays
non encore répertoriés, cela accroit le coût de 16,6%. La
langue la plus utilisée est l'espagnol, le français ensuite, l'arabe
pas du tout ! Il apparait que la littérature en anglais introduit des
biais dans tous les domaines, par exemple le coût en Europe est très
sous-évalué. Voir aussi Thiberge dans Le Monde du 31 mars 2021
: "les chercheurs ont épluché plus de 850 articles scientifiques...
près de 1 300 milliards de dollars (1 108 milliards d’euros) auraient
été perdus en l’espace de quarante ans... Mais ce chiffre
pourrait n’être que la partie immergée de l’iceberg.
« Ce coût global est très largement sous-estimé, explique
Franck Courchamp. Plus de 90 % des espèces envahissantes ne sont pas
encore évaluées. »"
150 espèces de fourmis ont été signalées hors de leur aire native et 30-35 sont installées de manière durable (Passera 2006). Passera parle d’espèces envahissantes, mais on utilisera l’anglicisme "invasives". Ce sont les « tramp species » ou fourmis vagabondes. On compte actuellement 241 espèces de fourmis introduites hors de leur zone native (Bertelsmeier et al 2017). Le laboratoire de Franck Courchamp travaille plus généralement sur les espèces invasives. "Quelles espèces, parmi la myriade de celles que comptent la faune et la flore, ont les caractéristiques requises pour être potentiellement envahissantes ? C'est tout le sens du travail de « profilage » auquel se sont livrés l'écologue et son équipe. Huit ans d'un labeur acharné, qui a abouti à une première publication, avant l'été, dans la prestigieuse revue « PNAS »" (Fournier et al 2019, Bertelsmeier et al 2013, Verdo 2019). Cleo Bertelsmeier, maintenant à Lausanne continue le travail sur les invasives.
En 2000, la liste des 100 organismes envahissants les plus dangereux
au monde établie par l’Union internationale de la conservation
de la nature (IUCN) comprenait 14 espèces d’insectes parmi lesquelles
5 espèces de fourmis (Lowe et al. 2000) : Anoplolepis gracilipes
(fourmi folle jaune, yellow
crazy ant), Linepithema humile (fourmi
d’Argentine, Argentine ant), Pheidole
megacephala (fourmi à grosse tête, big-headed ant), Solenopsis
invicta (fourmi de feu,
fire ant), Wasmannia auropunctata (the little fire ant, petite fourmi
de feu).
En 2005 on y ajoutait trois autres espèces (Passera and Aron 2005) :
Tapinoma melanocephalum (fourmi
fantôme, ghost ant), Monomorium pharaonis (fourmi
des pharaons) et Solenopsis geminata (fourmi de feu tropicale).
D’autres ont suivi depuis.
En 2015 l'IUCN déclarait 13 espèces de fourmis parmi 19 espèces
les plus invasives : Acromyrmex octospinosus, Anoplolepis gracilipes, Brachyponera
chinensis, Monomorium floricola, Monomorium pharaonis, Myrmica rubra, Paratrechina
longicornis, Pheidole megacephala, Solenopsis geminata, Solenopsis invicta,
Tapinoma melanocephalum, Technomyrmex albipes et Wasmannia auropunctata.
Toutes ces fourmis deviennt l'objet de commerce mondial comme animaux de compagnie
(Gippet et Bertelsmeier 2021).
En 2016 l'Union Européenne a défini une liste de 37 espèces
de plantes et animaux invasives contre lesquelles il faut lutter en prioirité,
mais pas de fourmis (Herzberg 2016b). Il ne faut pas oulier que d'autres insectes
sociaux sont invasifs comme des guêpes Polistes dominula (poliste
gaulois de nos régions, invasif en Amérique du Nord et Australie)
et Vespa velutina (frelon
asiatique), le termite de Formose Coptotermes
formosanus (de Taïwan, originaire de
Chine, arrivé en Louisiane avec les soldats à la fin de la deuxième
guerre mondiale). Même l'introduction de l'abeille domestique en Amérique
a eu des conséquences importantes sur les abeilles locales (Breed 2016).
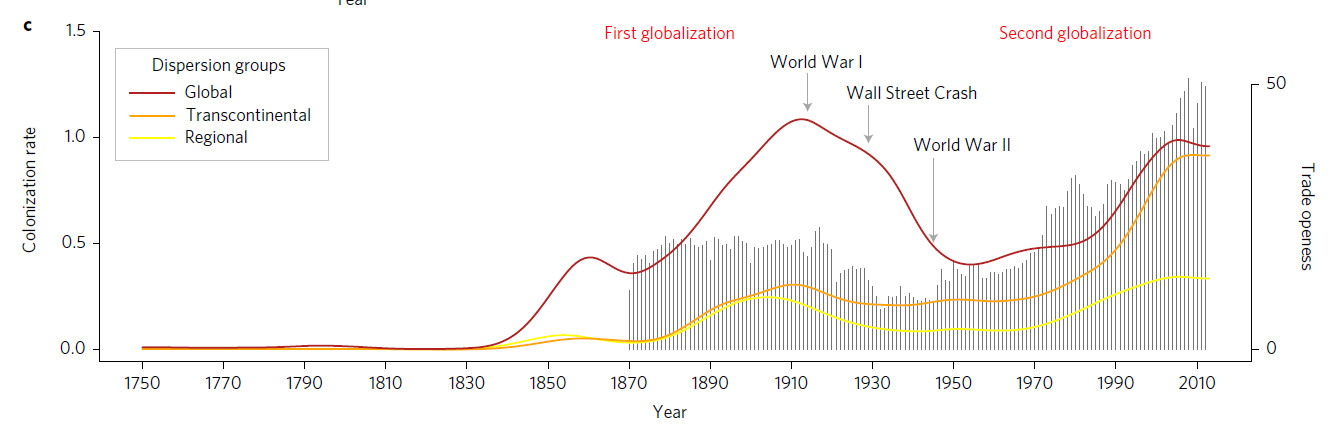
Dispersion des fourmis invasives. Un article important de Cléo Bertelsmeier à Lausanne a permis de montrer que les fourmis se sont dispersées sur le globe en suivant deux grandes vagues du commerce international, soit de 1850 à 1914 et de 1970 à nos jours (7sur7.be 2017, Affentranger 2017, Bertelsmeier et al 2017). On y apprend aussi qu'Acromyrmex octospinosus champignonniste de Guadeloupe considérée comme invasive ne l'est pas. Dans nos régions, les transports de fleurs en pot sont sans doute le principal moyen de dispersion. Selon Claude Lebas (mail du 4 novembre 2017) les vans pour chevaux seraient aussi une voie importante en Languedoc Roussillon pour Lasius neglectus. Selon Peter Wohlleben (2017, p.225) les palettes de bois insuffisamment chauffées pour être déparasitées offrent aux vagabonds de multiples opportunités pour traverser les océans. Le moustique tigre se déplace en co-voiturage depuis Barcelone dans toute l'Espagne selon Eritja et al (2017). Cleo Bertelsmeier a aussi fait une simulation de la progression prévisible des fourmis invasives en France avec le réchauffement climatique. Les ports d'entrée seraient surtout Biarritz, Toulon et Nice. La côte sud-ouest serait la plus touchée. Lasius neglectus et la fourmi d'Argentine auraient l'expansion la plus forte, suivies de Solenopsis richteri, Pheidole megacephala et Wasmannia auropunctata (Bertelsmeier and Courchamp 2014). Cleo Bertelsmeier a analysé des données collectées dans les ports maritimes et les aéroports des États-Unis et de Nouvelle-Zélande (Bertelsmeier et al 2018), elle déclare : "j’ai observé qu’une grande majorité des espèces transportées par l’homme ne proviennent pas de leur territoire d’origine, mais de régions précédemment colonisées. C’est notamment le cas dans 75% et 90% des fourmis entrant respectivement aux Etats-Unis et en Nouvelle-Zélande. Ces résultats laissent présager une augmentation des invasions biologiques sur le globe dans le futur, ce qui n’est pas très bon signe." (Mange 2018)
Les activités humaines transforment radicalement la circulation des êtres vivants sur la planète, y compris les microorganismes (bactéries, champignons, micro-algues) avec des répercussions probables sur les écosystèmes et les populations. On trouve par exemple une dispersion considérable de l'interféron de classe 1 qui favorise la résistance aux antibiotiques (Zhu et al 2017, voir Luk 2018).
Les fourmis invasives occupent des niches climatiques vides selon Arnan et al (2021). Une rechercche sur les bases de data sur 134 espèces fréquentes en Europe de l'ouest a été réalisée, 130 natives et 4 introduites (Lasius neglectus, Linepithema humile, Cardiocondyla emeryi et Pheidole megacephala). Le climat n'est pas le facteur déterminant pour les invasions, c'est plutôt les niches vides. Il est intéressant de noter que l'espèce la plus observée est Tapinoma cf nigerrimum qui correspond souvent à la nouvelle invasive T. magnum.
Turbelin (et al 2017 - voir aussi Courchamp 2021) ont analysé les bases de données de 2016 sur les espèces invasives animales et végétales dans divers pays nouvellement industrialisées comme Chine, l'Inde et le Brésil et des îles. Parmis les 1517 espèces, on trouve 39% des espèces qui ont été introduites volontairement, 29% involontairement et 22% les deux à la fois. En Nouvelle-Calédonie on trouve 183 invasives, à La Réunion 173. Le record est de 329 pour la Nouvelle-Zélande. Tapinoma melanocephalum est dans le top5 (les 25 plus invasives), elle est présente dans 40% des pays étudiés. Le commerce international de plantes ornementales est le principal responsable d'invasions de fourmis avec sous la rubrique "Horticulture" 116 espèces en Europe. Voir la revue de Xu et al (2022) sur les fourmis invasives en zone tropicale Asie-Pacifique, pays par pays, qui contient au moins 60 espèces invasives, y compris Solenopsis invicta, la plus destructive.
Les serres tropicales sont un milieu très favorable pour les fourmis invasives qui arrivent avec les plantes. Blatrix (et al 2018) en ont fait un inventaire en France et Belgique entre 2014 et 2016. Les oasis aussi accueillent des invasives, par exemple au Maroc (Taheri et al 2020)On trouve des Pheidole megacepha dans des serres
1
- Linepithema humile (fourmi d’Argentine, Argentine ant)
Elle est arrivée du sud du Brésil en 1915 (et pas d’Argentine)
sur la Côte d’Azur vers 1900 avec des plantes et s’est installée
depuis avec succès. Elle reste cantonnée le long des côtes
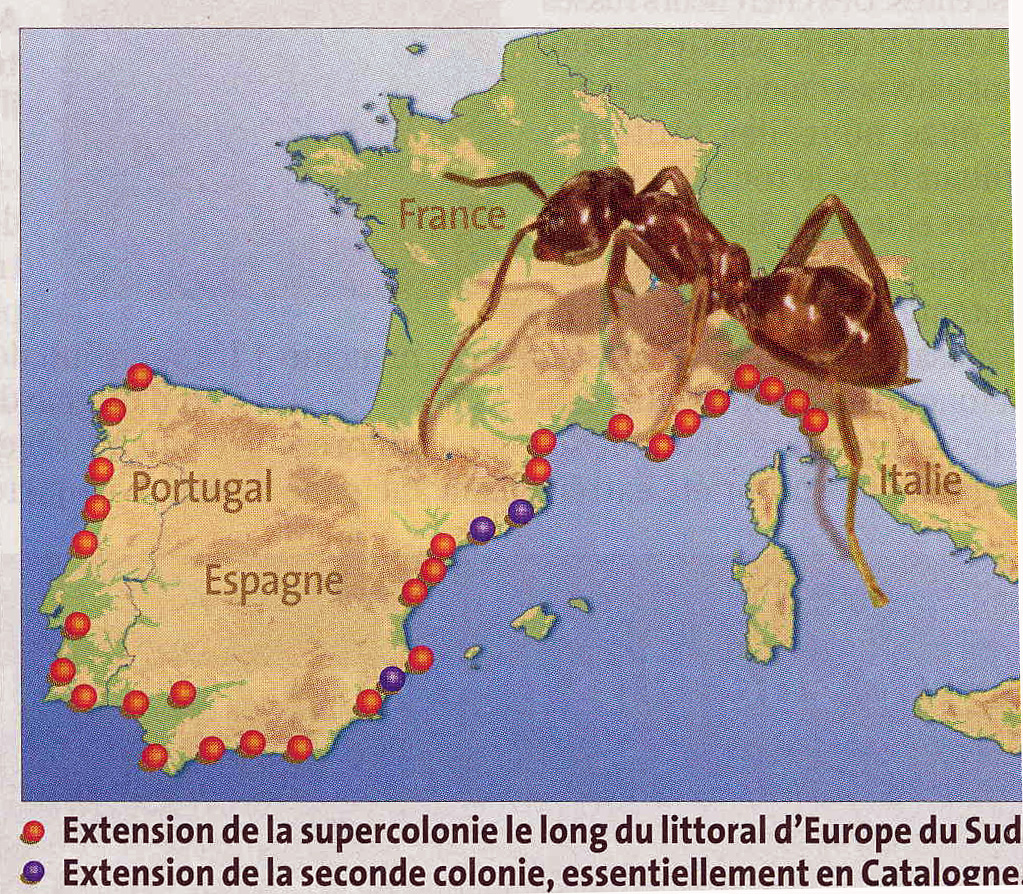
car elle ne supporte pas le gel. On a décrit deux super-colonies en Europe
le long de la côte méditerranéenne et jusqu’en atlantique
au Portugal (Giraud et al. 2002). En fait une nouvelle super-colonie a été
identifiée en Corse où le l’ai observée à
Calvi dans les années 70 (peut-être importée par les légionnaires
?) et au port des Oursinières dans une petite pinède à
l'Est de Toulon (Blight et al. 2010). C'est peut-être l'Argentine qui
perturbe les radars des autoroutes.. (Pontone 2014). En Corse on
attribue souvent les dégâts à la fourmi d'Argentine, mais
ce sont en fait des Tapinoma, les argentines n'étant que sur
la côte et elles semblent bien être les seules fourmis capables
de concurrencer les Linepithema (Chauvet 2018).

En septembre 2016 j'ai eu l'occasion de voir des pinèdes sur la côte au niveau de Girona entièrement envahies par des fourmis d'Argentine. C'est très impressionnant, il ne reste aucune autre fourmi. Paradoxalement, les fourmilions prospèrent dans cette zone, les fourmis étant abondantes. Mais cela ne suffit pas à limiter les argentines.
Pinède Escalas (près Girone) :



En 2016 elle a été trouvée à Nantes (Charrier et al 2020).
Andrew Suarez et son équipe ont examiné 40 sites déclarés infestés en Floride il y a 21 ans. La surface occupée a augmentée de 37% et la quantité de fourmis de 122% (avec des "pitfalls"). Le nombre d'espèces de fourmis natives sur ces site est passé de 14 à 4. En plus d'autres invasives sont apparues comme Brachymyrmex patagonicus et Pheidole flavens (Achury et al 2021)Pour en voir plus sur la fourmi d'Argentine
2
- Solenopsis invicta (fourmi de feu, fire ant)
C’est une véritable plaie aux USA où elle infeste plus de
140 millions d’hectares. Cela coûte 6,5 milliards de dollars par
an. Elle envahit maintenant l’Australie, Taïwan, la Chine, le Mexique
et les Caraïbes. Dans les zones infestées, 5% des personnes développent
une hyper-sensibilité au venin. Elle est connue pour former des radeaux
en cas d’inondation. Selon
Diagne (et al 2021) "Aux Etats-Unis, rien que les attaques des fourmis
de feu entraînent une centaine de milliers d’hospitalisations et
100 décès par an, pour un coût annuel de 6 milliards de
dollars."
En fait il y a plusieurs espèces comme S. richteri (Black Imported
Fire Ant – BIFA) longtemps considérée comme sous-espèce
de S. invicta. On la trouve au sud des USA, et serait originaire d’Amérique
Sud.
S. saevissima du Brésil est monocalique, mais en Guyane elle
forme des supercolonies (Martins et al. 2012; Dejean et al. 2015; Lenoir et
al. 2016).
S. geminata (Fourmi de feu tropicale) est dans tout le pacifique sud,
a sans doute été transportée depuis le Mexique par les
galions espagnols.

3 - Anoplolepis
gracilipes (Plagiolepidini, fourmi folle jaune, yellow crazy ant)
Cette fourmi est facile à reconnaitre à ses pattes très
longues et grêles, de grandes antennes et grands yeux et elle fait des
mouvements désordonnés très rapides d’où son
nom. Elle serait originaire d’Afrique et d’Asie. Elle est invasive
en Australie, Pacifique (Nouvelle-Calédonie, Hawaï, Galápagos,
Polynésie française), océan indien (Seychelles, île
Christmas, La Réunion). Elle est célèbre pour avoir détruit
plus de 20 millions de Crabes rouges terrestres (Gecarcoidea natalis)
endémiques de l'île Christmas (au Nord-Ouest de l'Australie et
au Sud-Ouest de Java) pendant leurs migrations annuelles. Voir
la revue de Lee et Scotty Yang (2022) sur cette fourmi qui se trouve du 27°N
au 27°S. C'est une revue complète sur
cette fourmi, la distribution, biologie, myrmécophiles, effets sur la
biodiversité, tous les essais de lutte y compris chimique.
Voir
plus


4 - Pheidole
megacephala (fourmi à grosse tête, big-headed ant, the Brown
House Ant, coastal brown ant en Australie)
On la trouve un peu partout en particulier dans les îles. En France dans
les régions parisienne (Muséum d'Histoire Naturelle de Paris). Dans
un zoo de serpents à Saint-Malo (Aligator Bay). A La Réunion pourrait avoir un
rôle positif de prédateur dans les plantations de manguiers - voir
plus loin (Jacquot et al 2016)

Jean Dorst signalait déjà cette invasion mondiale en 1965.
5 -
Wasmannia auropunctata (little fire-ant, petite fourmi de feu, fourmi électrique
en Nouvelle-Calédonie)
Wasmannia est
originaire d'Amérique tropicale, elle a envahit de nombreuses îles
du Pacifique y compris les Galapagos. En présence de Wasmannia
le nombre d'espèces de fourmis chute complètement, elle représente
90% à 99% des fourmis quand elle est présente. Elle monopolise
les sources de nourriture, il est possible que les autres espèces crèvent
de faim. En Israël il y aurait eu une seule invasion
(Vonshak et al 2009). Au
niveau européen, des nids de cette espèce invasive - originaire
d'Amérique centrale et du sud - ont été repérés
sous des serres en Angleterre (Geo
21 juillet 2022).
La fourmi électrique
a été détectée pour la première fois en France,
à Toulon en 2022. france3-regions.francetvinfo.fr
18 octobre 2022. Fourmi identifiée par Olivier
Blight.
Voir plus sur la petite fourmi de feu
6 - Tapinoma
melanocephalum (fourmi fantôme, ghost ant). Voir
Fourmi
fantôme
Fourmi tropicale toute petite facile à reconnaître avec son abdomen
clair que l’on peut colorer. Elle vit dans les maisons où elle
est d’une efficacité redoutable pour recruter des congénères
quand elles trouvent une source de nourriture.


La répartition de
la fourmi fantôme (Bertelsmeier et al 2017)
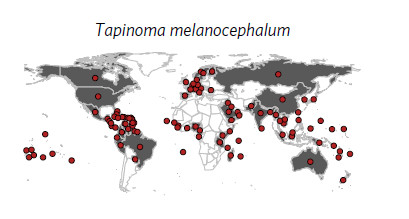
et pour voir la répartition actualisée (cabi.org). En France elle était connue des serres du Muséum d'histoire naturelle de Paris. J'en ai trouvé dans les serres tropicales du jardin botanique de Nancy en juin 2019. Elle est capable de mobiliser très vite un grand nombre d'ouvrières pour exploiter une nouvelle souce de nourriture (von Aesch et Cherix 2001).
7 - Monomorium
pharaonis (fourmi
des pharaons)
Originaire sans doute de la région éthiopienne, mais c’est
contesté. Elle vit dans les bâtiments bien chauffés des
grandes villes (par exemple dans les hôpitaux). En régressions
? À Tours elle était présente dans les années 70
dans divers immeubles, mais semble avoir disparue. Le laboratoire de Wilson
à Harvard a été envahi dans les années 60 par cette
fourmi (pdf)
Et plus récemment :
8 - Paratrechina longicornis
(Plagiolepidini) (Fourmi folle noire, longhorn (black) crazy ant)
C’est une fourmi surtout tropicale, qui peut s’installer jusqu’en
Estonie, Suède (habitations, serres). Origine Asie sud-est et Mélanésie
(Wetterer 2008). Trouvée en France en 1856 (Nylander), actuellement dans
des serres. Les reines isolées produisent des ouvrières diploïdes,
clones des reines, et des mâles clones de leur père (Pearcy et
al. 2011). C’est l’alliée de Ant-man, le film (2015). Voir
Fourmis folles

9 - La fourmi folle rousse
/ fauve Nylanderia fulva (Rasberry crazy ant, tawny crazy ant)
C’est une fourmi originaire d’Argentine et du sud du Brésil
qui semble contrarier l’expansion des fourmis de feu aux États-Unis
(arrivée en 2002 au Texas). Elle est rousse parsemée de poils
rouges et fait de gros dégâts aux matériels électriques.
Elle est en expansion rapide et pourrait provoquer plus de dégâts
que la fourmi de feu. Voir Fourmis
folles
10 -
En Europe, Lasius neglectus (Invasive garden ant, super fourmi asiatique,
fourmi
envahissante des jardins, fourmi
aztèque, voir plus)
C’est la fourmi de la Mer Noire selon Fraval (Fraval 2009).
Elle est originaire des steppes d’Asie mineure, sans doute de Lasius
turcicus qui a deux formes, d’altitude unicoloniale et de plaine
supercoloniale (Cremer et al. 2008; Ugelvig et al. 2008,
Ugelvig and Cremer 2012). Les colonies comportent
de nombreuses
reines de petite taille avec accouplement intranidal (pas de vol nuptial), elles
forment des super colonies (jusqu’à 35 000 ouvrières) avec
de nombreuses ouvrières « nanitic » de petite taille qui
se développent plus vite (Espadaler
et Rey 2001).
On la trouve beaucoup dans la région lyonnaise (Gippet et al 2021).
À signaler sa progression rapide en France, en Touraine
on la trouve dans les jardins de La Riche depuis les années 2000. J'en
ai reçu récemment de Montlouis sur Loire, de Saint-Brévin
où elle semble détruire les nids de Messor. Elle a envahi
le village de Désirat en Ardèche (Plummer 2015). Elle
peut faire des dommages
aux circuits électriques. On la trouve beaucoup
dans la région lyonnaise (Gippet,
thèse 2016). Elle se déplace avec les transports de terre, béton
lors de travaux (Gippet et al 2017).
Voir tout sur la répartition et la biologie sur le site du CREAF
à Barcelone (Espadaler and Bernal 2015). Son expansion pourrait être
ralentie par un champignon Laboulbenia formicarum (Tragust et al. 2015),
mais je n’y crois pas trop car l'immunité
sociale est très forte chez les fourmis. Lasius neglectus
est arrivée en Angleterre en 2010. Elle est adaptée aux climats
plus frais, par exemple à 30°C elle ne résiste pas bien et
est combattue efficacement par les Crematogaster et Tapinoma
(Frizzi et al 2017).


11 - Myrmica
rubra (Fourmi rouge, ruby ant) d’Europe
Notre fourmi rouge est connue depuis 1900 au Massachussetts (Wheeler 1908).
Elle semble être devenue invasive aux USA dans le Maine dans les années
50 et actuellement dans 4 autres états (Groden et al. 2005), mais reste
multicoloniale (Garnas 2007). On la trouve aussi à Terre-Neuve et au
Canada. À Terre-Neuve, avec des analyses génétiques, il
y a au moins 4 sources d’importation possibles (Hicks et al. 2014). Pour
le bilan de la répartition en Amérique du Nord voir (Wetterer
and Radchenko 2011). On l'appelle maintenant "European
Fire ant". Les odeurs de M. rubra font peur aux araignées
locales au Canada (voir Araignées).

12
- D'autres espèces
- Diverses espèces deviennent invasives en milieu tropical ou subtropical,
par exemple Ooceraea
(Cerapachys) biroi (fourmi maraudeuse, clonal raider ant) dans les
îles ( Wetterer et al 2012).
- J'ai trouvé dans mon village à Sauvagnon (64230) en avril 2017 Tapinoma magnum, une fourmi originaire de la côte méditerranéenne de Narbonne à Montpellier, en Corse et en Afrique du Nord. Elle a été trouvée aussi dans la banlieue de Bordeaux et elle est signalée comme invasive en Allemagne, Belgique (Ostende) et Pays-Bas. Elle arrive sans doute avec des plants. Voir plus sur T. magnum
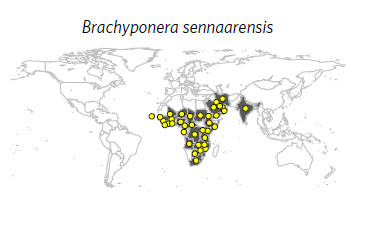
- Brachyponera sennaarensis (samsun ant) selon Bertelsmeier et al 2017 :
Brachyponera chinensis (Asian needle ant, fourmi aiguille asiatique) a colonisé les USA dans les années 1930. Elle devient invasive dans les états du sud depuis les années 2000 où elle fait reculer la fourmi d'Argentine. C'est une fourmi qui se nourrit de termites Reticulitermes flavipes. Pour lutter contre cette fourmi on teste de leur donner des termites empoisonnés avec du fipronil. Ce serait plus efficace que des appâts avec "moins" de pollution environnementale... bof .. (Buczkowski 2016). Aux USA on a étudié la progression de cette fourmi dans les forêts apalachiennes où elle est en concurrence acec Aphaenogaster rudis et A. picea. Hélas les seuls refuges pour les deux espèces locales sont l'altitude et les zones plus nordiques où B. chinensis ne supporte pas le froid. Peu encourageant pour la lutte contre les invasives !

- Tapinoma sessile, la fourmi odorante, d'Amérique du Nord est aussi envahissante dans certaines zones.
- Tetramorium bicarinatum, originaire d'Asie du Sud-Est, introduite un peu partout en pays tropicaux et en France dans les serres tropicales. Cyril Astruc, pour sa thèse a effectué des tests d'agression entre fourmis originaires du Japon (locales) et du Brésil (invasives) et n'a pas observé d'agressivité entre colonies (unicolonialité) alors qu'elles sont très agressives envers d'autres espèces.
Parmi les fourmis vendues dans le monde il y a 57 invasives, reconnues comme les plus invasives au niveau mondial par l'IUCN : Acromyrmex octospinosus, Anoplolepis gracilipes, Brachyponera chinensis, Monomorium floricola, Monomorium pharaonis, Myrmica rubra, Paratrechina longicornis, Pheidole megacephala, Solenopsis geminata, Solenopsis invicta, Tapinoma melanocephalum, Technomyrmex albipes et Wasmannia auropunctata (Gippet et Bertelsmeier 2021).
13
- Le cas des îles
Pour plus d'infos sur les envahisseurs (plantes
et animaux) voir le livre de Albouy
(2017). On y trouve par exemple le cas de l'île de la Surprise (24
ha) en Nouvelle-Calédonie où les rats qui faisaient des ravages
dans les populations d'oiseaux et de reptiles ont été éliminés
avec succès sans que cela perturbe la population de fourmis Pdf.
Selon Cerdá et al (2012) il y avait sur cette île une seule espèce
locale, Pheidole oceanica, qui résiste face à 8 espèces
introduites. Voir aussi plus haut le cas de la fourmi maraudeuse
Ooceraea (Cerapachys)
biroi qui devient invasive dans les îles. Technomyrmex
brunneus (Dolichoderinae), Fourmi
d'Asie de sud a envahi de nombreuses îles au Japon, Corée, Bornéo
et Nouvelles-Guinée (Pitri et al 2021). A
l'île de La Réunion Fabrice
Blard signalait déjà en 2003 que la moitié des 27 espèces
trouvées sont "vagabondes" (Pdf)
Effectivement, les îles sont les premières victimes des espèces invasives. Parmi les 1288 îles accueillant des vertébrés menacés, 1030 abritent également des espèces invasives (Garric 2017).
Caractéristiques
des invasives
Toutes ces «tramp species » ont des caractéristiques communes
(Passera 1993 version française, 1994) (voir Passera and Aron 2005 -
chapitre 15 et Passera 2006 pages 217 et suiv.) :
- propension à envahir de nouveaux biotopes et en particulier les îles,
- unicolonialité (formant des supercolonies sans agression). C’est
ainsi que Tapinoma sessile en Indiana change de statut avec l’urbanisation
: en milieu naturel elle forme des colonies simples monogynes, monocaliques,
et elle devient polygyne, supercoloniale et envahissante en milieu semi-naturel
et urbanisé où elle fait disparaître les autres espèces
(Buczkowski 2010). Lepisiota canescens
native d'Ethiopie est unicoloniale et semble être une invasive potentielle
qui a déjà envahi l'Afrique du Sud et est arrivée à
Darwin en Australie. Il n'y aurait qu'une vingtaine d'espèces unicoloniales.
- commensales de l’homme,
- très polygynes avec des reines à durée de vie très
courte s’accouplant dans le nid sans vol nuptial,
- multiplication par bouturage,
- et enfin ouvrières toujours stériles.
Pourtant, selon Passera, un seul caractère leur est propre : la dispersion
par les activités humaines.
Selon Bertelsmeier
(et al 2017) les espèces de petite taille et dont les colonies
comportent plusieurs reines sont plus enclines à se répandre.
En conséquence il y a perte de la diversité génétique
et les flux de gènes entre supercolonies sont faibles. C’est ainsi
que les Wasmannia d’Israël proviennent d’une seule
reine d’Amérique du sud (Hefetz). Ces fourmis changent facilement
de profil d’hydrocarbures cuticulaires «You are what you eat»
comme cela a été montré chez Linepithema qui intègre
des alcanes des grillons donnés au labo (Liang and Silverman 2000). On
vient de montrer aussi que le frelon asiatique invasif en France, malgré
une grande homogénéité génétique, présente
des variations de profils cuticulaires, indiquant une grande flexibilité
(Gévar et al 2017).
Tapinoma
sessile,
une fourmi native des USA, où elle forme des petites colonies monocaliques
dans un seul nid avec une seule reine, et qui devient invasive en zones urbanisées
(polydomie, polycalisme). La stucture génétique est extrêmement
plastique pour cette espèce. L'urbanisation entraîne une restriction
du flux de gènes et une forte pression de sélection (Blumenfeld
et al 2021).
Héritabité des hydrocarbures : chez Monomorium pharaonis en élevage au laboratoire les hydrocarbures ont peu d'héritabilité, ce qui semble indiquer qu'ils sont sujets à sélection. C'est sans doute ce qui explique qu'en élevage en laboratoire ils varient facilement, cela pourrait etre une caractéristique des fourmis invasives (Walsh et al 2019). Ce ne serait pas le cas de Tapinoma magnum qui garde longtemps en laboratoire son profil d'hydrocarbures (Lenoir et al 2022).
D’une manière générale,
les fourmis invasives peuvent perturber les mutualismes, voir la synthèse
de Kiers et al. 2010.
Les invasions biologiques s'accompagnent aussi de modifications
du microbiote intestinal comme c'est le cas pour le termite Reticulitermes
flavipes originaire de Louisianne et introduit en France au cours du 18ème
siècle (Dedeine et al 2017).
Pourtant bonne nouvelle
selon Cleo
Bertelsmeier : Le
Journal du Jura (2020) Les espèces invasives les plus dangereuses sont
les plus casanières. journaldujura.ch
15 octobre 2020.
"Cleo Bertelsmeier, professeure assistante au Département d’écologie
et évolution de l’UNIL, et sa doctorante Olivia Bates ont voulu
savoir si les espèces introduites les plus invasives sont les plus aptes
à coloniser de nouveaux climats. Pour ce faire, les biologistes ont évalué,
en collaboration avec Sébastien Ollier, du Laboratoire Ecologie, Systématique,
Evolution de l’Université Paris-Saclay, la niche climatique de
82 espèces de fourmis provenant du monde entier. Résultats: "Nous
avons pu démontrer que les espèces les plus néfastes pour
l'être humain et l’environnement s’aventurent paradoxalement
le moins dans des climats auxquels elles ne sont pas exposées dans leur
zone native. En d’autres termes, les espèces les plus dangereuses
sont plus frileuses et moins expansionnistes que les autres", commente
Cleo Bertelsmeier, citée dans le communiqué. Ces résultats
constituent une bonne nouvelle, comme le relève Olivia Bates, première
auteure de l'article: "Puisque les espèces invasives les pires sont
peu enclines à coloniser de nouveaux climats, leur propagation spatiale
est prévisible. Il suffit de se baser sur l’ensemble des conditions
climatiques dans leur aire native pour élaborer un modèle prédictif
qui identifie les endroits sur Terre présentant des conditions climatiques
similaires, propices à l’espèce".
(Bates et al 2020)
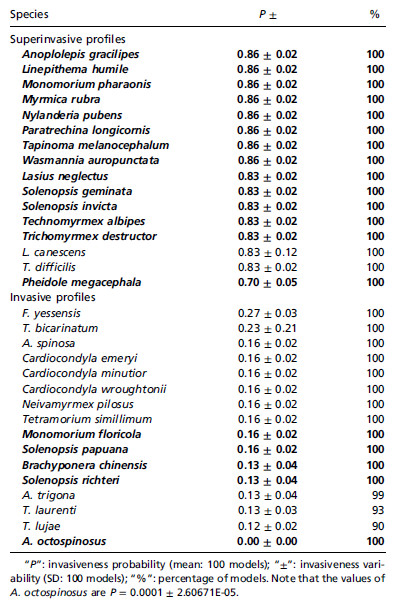
Pourquoi ces fourmis deviennent-elles invasives ? C'est une question sans véritable réponse pour le moment. Calcatera et al (2016) ont étudié Solenopsis richteri et Linepithema humile dans leur région native en Argentine. Les deux espèces cohabitent avec 47 autres espèces sans problèmes. Les auteurs émettent l'idée que ces fourmis sont écologiquement flexibles... C'est peut-être lié aux changements de milieu comme Tapinoma sessile en Indiana qui change de statut avec l'urbanisation (voir plus haut Buczkowski 2010). Pourtant, selon l'équipe de Franck Courchamp, il est possible de prédire la probabilité que certaines fourmis soient invasives selon leur profil écologique et leur attribuer le caractère de "profiler" (Fournier et al 2019. Voir Courchamp 2019 avec la liste des fourmis super-invasives (probabilité >80%) et des invasives potentielles (10 à 30%) Liste (Acromyrmex octospinosus de Guadeloupe avec 0% est éliminée de la liste des invasives)
Le succès de T. immigrans (espèce cryptique) face à T. caespitum serait lié à des accouplements multiples en particulier avec des mâles de l'autre espèce. Voir une synthèse sur le genre Tetramorium en France de M. Cordonnier.Des mâles
plus gros et plus efficaces chez T. immigrans :
Vol nuptial et polyandrie interspécifique chez les deux espèces :
Marie-Monique Robin dans son livre La fabrique des pandémies (2021) émet une hypothèse à propos des espèces invasives : "Plusieurs études suggèrent que les rongeurs généralistes ont une certaine plasticité génétique, avec des récepteurs capables de se lier à de nombreuses maladies, mais aussi de les transmettre. Ils sont aussi associés à des environnements perturbés, peut-être du fait d'un avantage évolutif qui leur permet de s'adapter à des habitats variés et de vivre à proxilmité des humains.". Est-ce que les fourmis invasives seraient aussi plus plastiques génétiquement ?
Compétition
entre invasives
Tapinoma nigerrimum semble combattre efficacement la fourmi d’Argentine
(Blight et al. 2010), mais elle devient aussi une peste.. On la trouve dans
les bouches du métro à Lyon et même en Belgique. T.
nigerrimum est souvent considérée comme nuisible mais elle
mange les larves de la mouche des fruits Ceratitis capitata dans les
vergers de citronniers en Italie (Calabre). Les autres fourmis (Messor structor,
Camponotus aethiops et Formica fusca) très peu efficaces
contre l'Argentine (Campolo et al. 2015). La
compétition Tapinoma magnum / fourmi d'Argentine est réelle.
Christain Foin me dit "avec Claude [Lebas]
nous avons vu un affrontement entre deux énormes colonies de Linepithema
et de Tapinoma. J’y aurais passé la journée à les
regarder. C’était une guerre de positions et une guerre chimique.
Deux armées d’une quinzaine de cm de large face à face avec
un no man’s land entre les deux." (Christian
Foin, mail du 9 novembre 2019).
Cleo Bertelsmeier dans le laboratoire de Franck Courchamp à Orsay a travaillé sur la compétition entre 4 espèces de fourmis invasives en les testant en laboratoire par des combats entre deux espèces. Les espèces utilisant des armes chimiques (Wasmannia auropunctata et Lasius neglectus) sont bien meilleures que les autres qui utilisent leurs simples mandibules (Pheidole megacephala qui perd toujours en combat à deux et Linepithema humile). P. megacephala a cependant des ressources puisque en combat à quatre elle laisse les autres s'entretuer pour prendre la place ensuite (Stratégie de Napoléon selon Herzberg) (Bertelsmeier et al. 2016). Voir l'article du Monde (Herzberg 2016a Pdf), Le Journal du Dimanche (Bellet 2016 Pdf) et Le Figaro (Nothias 2016 Pdf).
Compétition entre la fourmi d'Argentine et la fourmi aztèque Lasius neglectus. Ces deux espèces invasives sont amenées à se retrouver de plus en plus souvent en compétion en Europe. Des chercheurs espagnols et polonais ont étudié en laboratoire cette compétition. Elles utilisent deux stratégies connues chez les fourmis : la stratégie du bourgeois, où l'on change d'agressivité en fonction de la dominance numérique, et la stratégie du cher-ennemi où l'on accepte l'autre avec territoire partagé (Trigos-Peral et al 2020). Des préjugés
Attention aux idées préconçues et non vérifiées
: les invasives font disparaître les fourmis natives comme à Port-Leucate
où ne subsistent que 2 espèces devenues très rares (Passera
2006). Cependant, à Madère (Açores) Pheidole megacephala
puis Linepithema humile sont censées avoir éradiqué
toutes les espèces natives. En fait, c’est faux, ces deux espèces
occupent 0,3 et 6% de l’île, et n’ont jamais occupé
plus de 10%. Sur 10 espèces natives, 9 sont encore présentes (Wetterer
2006). Aux Galapagos la communauté des fourmis comporte 29 espèces
dont 22 introduites ; surtout S geminata et W auropunctata. Mais les
fourmis natives survivent encore !! (Peck et al. 1998).
Vive
les envahisseurs ?
C'est
un peu iconoclaste mais parfois on peut se demander si les envahisseurs ne sont
pas une solution à la destruction d'écosystèmes perturbés.
C'est ainsi qu'à Hawaï un oiseau envahisseur aurait plutôt
un rôle positif dans la dispersion des graines... (Herzberg 2019).
Il faut sans doute
s'inspirer de ce que dit le philosophe Baptiste Morizot "On a longtemps
cru qu'il fallait exploiter plus efficacement la nature pour améliorer
nos conditions de vie humaine; on commence à comprendre, que pour atteindre
cet objectif, il faut apprendre à mieux cohabiter avec les autres créatures
de la terre." (Vincent 2016)
Invasives et lutte
biologique
Les fourmis invasives peuvent même avoir un rôle dans la lutte biologique.
Cela a été étudié à La Réunion dans
des plantations de mangues où Pheidole megacephala et Solenopsis
geminata mangent beaucoup d'oeufs de la mouche des fruits Zeugodacus
cucurbitae (Jacquot et al 2016). Ces mêmes P. megacephala
sont aussi de bons agents prédateurs des larves du charançon du
café Xylosandrus compactus à La Réunion (Ogogol
et al 2017).
Des
invasives qui disparaissent
Des fourmis invasives peuvent disparaitre spontanément
sans intervention humaine, c'est ce qu'ont observé M Cooling et B Hoffmann
en Australie sur la fourmi folle jaune (yellow crazy ant, Anoplolepis gracilipes)
entre 2003 et 2014, une invasive qui pose beaucoup de problèmes (Cooling
and Hoffmann 2015). Cette disparition pourrait être due à des virus,
et d'autres bactéries pathogènes (Cooling et al 2016).
Intéressant, cela me rappelle une observation de Luc Passera qui signalait
que la fourmi d'Argentine semble régresser spontanément sur la
côte méditerranéenne vers Leucate. La fourmi d’Argentine
qui avait envahi la Nouvelle-Zélande semble régresser spontanément
(Cooling et al. 2012).
A signaler un joli travail de l'équipe d'E. Darrouzet avec S. Aron (Bruxelles) sur la dépression de consanguinité des frelons asiatiques arrivés en France lors d'une seule introduction. Cela aboutit à donner des mâles diploïdes stériles (Darrouzet et al 2015). Est-ce que cela aboutira à une limitation de l'invasion ? A mon avis peu probable, mais ce serait mieux que je me trompe. D'ailleurs je suis très surpris qu'on n'ait pas encore signalé d'autres arrivées..
Le coût des espèces de fourmis invasives (Angulo et al. 2022). Une analyse des coûts sur 1342 rapports dans 27 pays pour 12 espèces de fourmis depuis 1930 arrive à 52 milliards de dollards. 80% de ce total provient de deux espèces emblématiques : Solenopsis invicta (RIFA) et Wasmannia auropunctata. Autres espèces étudiées : Anoplolepis gracilipes, Linepithema humile, Acromyrmex octospinosus, Lepisiota frauenfeldi, Pheidole megacephala, Lasius neglectus, Trichomyrmex destructor et Monomorium pharaonis. Cette estimation est sans doute très sous-évaluée car les données pour les pays en voie de développement sont mal connues.
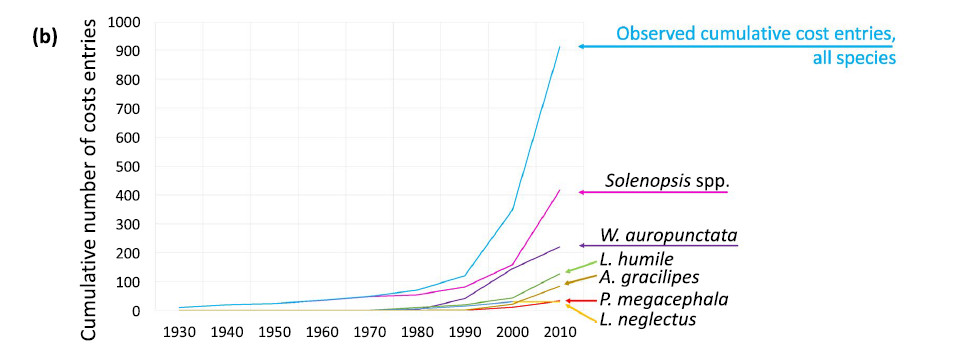
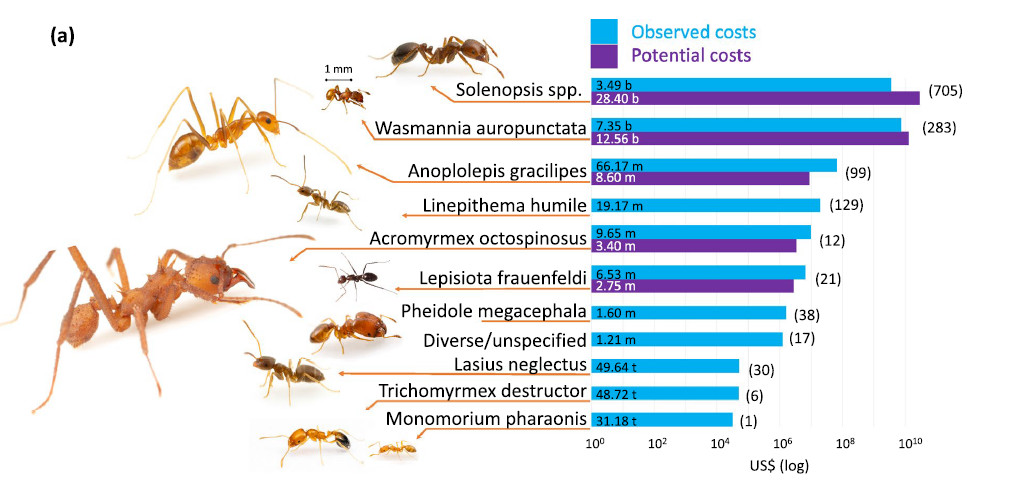
Le
vrai coût des fourmis envahissantes révélé par des
chercheurs marocains (bladi.net
2 juin2022)
"Une étude internationale menée par le Conseil supérieur
de la recherche scientifique (CSIC), et à laquelle ont participé
des chercheurs marocains, a révélé que les invasions des
fourmis envahissantes ont eu un impact économique négatif sur
l’agriculture et la santé publique depuis 1930, entrainant des
dépenses estimées à 46 milliards d’euros.
Les fourmis envahissantes perturbent les écosystèmes et sont responsables
de la disparition d’autres espèces de fourmis. Mais elles ont également
impacté, d’un point de vue économique, l’agriculture
et la santé publique depuis 1930, entraînant des dépenses
estimées à 46 milliards d’euros. C’est la conclusion
de l’étude internationale menée par le CSIC et publiée
dans la revue Biological Invasions. Des chercheurs du Maroc, d’Australie,
de France, d’Italie, de République tchèque, d’Inde,
du Koweït et du Japon ont participé à ces travaux, rapporte
Sabemos.
« Les fourmis envahissantes peuvent être très dangereuses
pour la santé humaine et pour les animaux de la ferme. […] Son
impact peut être très néfaste pour les cultures »,
explique Elena Angulo, chercheur au Centre biologique de Doñana (EBD-CSIC).
Elles perturbent les chaînes alimentaires, diminuent la pollinisation,
affectent la production agricole, endommagent les infrastructures et peuvent
mettre en danger la santé publique, ajoute-t-elle.
L’étude s’est concentrée sur l’analyse des informations
relatives à 12 des 19 espèces de fourmis identifiées comme
envahissantes par l’Union internationale pour la conservation de la nature
(UICN). Sur les 46 milliards d’euros, quelque 9,4 milliards ont été
investis dans la gestion des invasions et un montant plus faible dans la mise
en place de mesures préventives. Toutefois, « beaucoup de frais
de recherche n’ont pas été comptabilisés, car les
administrations ne les signalent pas », souligne Elena Angulo, invitant
à « une plus grande collaboration » entre les différents
acteurs afin de déterminer « le véritable impact de ces
invasions et améliorer les stratégies de gestion ». "
(Angulo et al 2022)
Lutte
contre les fourmis invasives
Dans les années 1950 aux USA avec l'apparition de nouveaux insecticides,
une grande campagne d'éradication de la fourmi de feu a été
lancée sur 56 millions d’hectares. Cela a été un
échec cuisant. Voir
la critique de Rachel
Carson dans son livre "Le printemps silencieux" (Carson 1968).
E. O. Wilson, le célèbre myrmécologue américain,
a d’ailleurs qualifié cette guerre perdue de « Vietnam entomologique
» (Wilson 200, p. 236, voir Keller et Gordon 2006 p. 188). Des essais
de lutte biologique sont menés depuis des années contre Solenopsis
invicta par des champignons, des parasitoïdes (Eucharitidae, mouches
phorides) qui n’ont pas suivi lors de la migration. On peut infecter les
fourmis avec le champignon pathogène Metarhizium anisopliae,
mais elles boivent plus de quinine (self medication), reçoivent plus
de trophallaxies (Qiu et al. 2016). C’est de l’immunité sociale,
donc la lutte est difficile. Pseudacteon tricuspis et P. curvatus
sont des mouches phorides parasitoïdes d'Amérique du Sud qui pourraient
limiter la prolifération des fourmis de feu, mais leur effet est minime
(Valles et al. 2010; Porter and Calcaterra 2013), même si des essais continuent
(Porter et al 2018). Pierre Jolivet écrivait déjà en 1986
que la lutte avec les parasitoïdes « semble sans espoir
» (Jolivet 1986, p. 187).
En Nouvelle-Calédonie, il est sans doute déjà trop tard
pour stopper l'invasion (tahiti.info 2017). Attention aux effets pervers : en
Nouvelle-Zélande, la fourmi
d'Argentine fait disparaître les Monomorium antarcticum locales.
L'imidaclopride abaisse l'agressivité des Monomorium et au contraire
augmente celle des Argentines, ce qui augmente leur chance de se répandre
(Barbieri et al 2013, voir Bossy 2013).
Hoffmann a fait le bilan de 316 campagnes d’éradication
pour 11 espèces d’invasives, hélas avec des insecticides
puissants comme le fipronil (qui pose des problèmes pour les abeilles
!), l’hydraméthylnone (pour cafards et fourmis), des régulateurs
de croissance (pyriproxyfène et méthoprène). Il note 144
succès surtout en Australie mais uniquement sur très petites surfaces
(Hoffmann et al. 2016).
Des essais de lutte sont conduits actuellement avec des produits comme des extraits
de thé, soit les feuilles soit les restes après usage, contenant
diverses molécules toxiques comme des polyphénols sur Paratrechina
longicornis, Anoplolepis gracilipes (fourmis
folles) en Malaise (Dieng et al. 2016).
Contre Tapinoma
melanocephalum (fourmi fantôme), la fourmi d’Argentine
et la fourmi des pharaons à Taiwan on peut utiliser des solutions avec
de l’acide borique à 1% et du sucre, efficace en 4 semaines (Klotz
et al. 1996; Luo and Chang 2013).
et si on laissait faire la nature comme le dit Vincent Albouy
? 
Selon Cleo Bertelsmeier : "Nous sommes donc loin d'en avoir fini avec les fourmis invasives - des espèces qui doivent leur formidable succès écologiques à leur organisation sociale et à différentes stratégies comportementales de dominance physique et de monopolisation des ressources." (Les guerres secrètes des fourmis, p.193).
C'est aussi ce que dit Quentin Rome à propos du frelon asiatique "On ne pourra très probablement pas éradiquer ou juguler l'extension du frelon asiatique. D'ailleurs, on y est parvenu pour aucune guêpe sociale. Mieux vaudrait trouver une méthode pour vivre avec." Pdf
Comment les fourmis voyagent ? Dans Minuscules 2 on a une réponse : les fourmis rouges se retrouvent dans un carton en direction de Tokyo où elles vont bien sûr s'installer.
Remerciements. Merci à Laurence Berville pour avoir envoyé des fourmis invasives d'Australie (dans l'alcool bien sûr).
Références
- 7sur7.be (2017) Comment les fourmis ont conquis le globe. 7sur7.be, 22 juin
2017, p. http://www.7sur7.be/7s7/fr/1505/Monde/article/detail/3189541/2017/06/22/Comment-les-fourmis-ont-conquis-le-globe.dhtml
Pdf
- Affentranger, M. (2017) Des fourmis dans nos bagages. unil.ch, 22 juin 2017,
p. http://www.unil.ch/fbm/home/news.html?showActu=1498140017882&showFrom=1.
Pdf
- Angulo,
E., C. Diagne, L. Ballesteros-Mejia, T. Adamjy, D. A. Ahmed, E. Akulov, A. K.
Banerjee, C. Capinha, C. A. K. M. Dia, G. Dobigny, V. G. Duboscq-Carra, M. Golivets,
P. J. Haubrock, G. Heringer, N. Kirichenko, M. Kourantidou, C. Liu, M. A. Nuñez,
D. Renault, D. Roiz, A. Taheri, L. N. H. Verbrugge, Y. Watari, W. Xiong and
F. Courchamp (2021). Non-English languages enrich scientific knowledge: The
example of economic costs of biological invasions. Science of The Total Environment:
144441. doi: https://doi.org/10.1016/j.scitotenv.2020.144441. Pdf
-
Angulo, E., B. D. Hoffmann, L. Ballesteros-Mejia, A. Taheri, P. Balzani, A.
Bang, D. Renault, M. Cordonnier, C. Bellard, C. Diagne, D. A. Ahmed, Y. Watari
and F. Courchamp (2022). Economic costs of invasive alien ants worldwide. Biological
Invasions: 1-20. doi: 10.1007/s10530-022-02791-w. Libre
de droits
- Bellet, R. (2016). Et maintenant, la fourmi électrique. Le Journal
du Dimanche 30 avril 2016. Pdf
-
Blumenfeld, A. J., P.-A. Eyer, A. M. Helms, G. Buczkowski and E. L. Vargo (2021).
Consistent signatures of urban adaptation in a native, urban invader ant Tapinoma
sessile. Molecular Ecology in press. doi: https://doi.org/10.1101/2021.06.21.449338
- Bossy, D. (2013) Les
néonicotinoïdes poussent les fourmis à se battre jusqu'à
la mort. Futura Science, 25 octobre 2013, p. http://www.futura-sciences.com/planete/actualites/developpement-durable-neonicotinoides-poussent-fourmis-battre-jusqua-mort-49809/
Pdf
-
Chauvet, A. (2018) Chaleur, humidité, déchets : l'été
rêvé des fourmis invasives. corsematin.com, 31 août 2018.
https://www.corsematin.com/article/article/chaleur-humidite-dechets-lete-reve-des-fourmis-invasives.
Avec interview d'Alain Lenoir. Pdf
-
Cordonnier, M., B. Kaufman and G. Escarguel (2020) S’accoupler plus souvent,
et avec des mâles d’autres espèces : la clé du succès
d’une invasion biologique chez les fourmis ? theconversation.com
27 octobre 2020.
-
Courchamp, F. (2019) Des chercheurs mettent au point une méthode pour
détecter les espèces envahissantes 3 avril 2019, p. https://theconversation.com/des-chercheurs-mettent-au-point-une-methode-pour-detecter-les-especes-envahissantes-114726Autor
. Pdf
-
Eyer, P.-A., P. T. Shults, M. R. Chura, M. N. Moran, M. N. Thompson, A. M. Helms,
R. K. Saran and E. L. Vargo (2021). Divide and conquer: Multicolonial structure,
nestmate recognition, and antagonistic behaviors in dense populations of the
invasive ant Brachymyrmex patagonicus. Ecology and Evolution n/a(n/a). doi:
https://doi.org/10.1002/ece3.7396.
- Fraval, A. (2009). La fourmi
noire de la Mer noire. Une invasion bien préparée. Insectes 152:
32. Pdf
- Garric, A. (2017). Les îles sont des épicentres du déclin
de la biodiversité. lemonde.fr 26
octobre 2017. http://www.lemonde.fr/biodiversite/article/2017/10/26/comment-sauver-40-des-especes-de-vertebres-en-danger-critique-d-extinction_5206262_1652692.html#HxUqyR7qfr6t4qkk.99.
Pdf
et Le Monde 1-2 novembre 1017. p. 6. Pdf
- Herzberg, N. (2016a). La
guerre des fourmis. Le Monde Science et Médecine 2 mai 2016. Pdf
-
Herzberg, N. (2019). Les envahisseurs et la biodiversité. Le Monde 17
avril 2019. Pdf
- Lenoir, A., S. Devers,
A. Touchard and A. Dejean (2016). The Guianese population of the fire ant Solenopsis
saevissima is unicolonial. Insect Science in press. Doi: 10.1111/1744-7917.12232.
Pdf
-
Luk, F. L. (2018) Le
tour du monde des microbes. lejournal.cnrs.fr, 11 janvier 2018
- Mange, F. (2018) Des fourmis qui voyagent toujours plus loin. letemps.ch,
7 mai 2018, https://www.letemps.ch/sciences/fourmis-voyagent-toujours-plus-loin.
Pdf
- Normand,
J.-M. (2019). L'inexorable invasion du frelon asiatique en France. Le Monde
15 juin 2019. p. 6. Pdf
- Nothias, J.-L. (2016).
Les fourmis sont des guerrières hors pair. Le Figaro Mercredi 20 avril
2016. Pdf
- Passera, L. (1993). Quels sont les caractères étho-phusiologqiues
des 'Fourmis vagabondes"? . Actes Coll. Insectes Sociaux 8: 39-45. Pdf
-
Pham, Q. (2021) Les espèces invasives, fléau méconnu qui
coûte des milliards. huffingtonpost.fr
31 mars 2021.
- Plummer, W. (2015). Ardèche
: un village envahi par des fourmis. Le Figaro 8 avril. Pdf
- Pontone, C. (2014). Toulon : le radar de l'A57 refuge d'une fourmilière.
Var Matin 3 Octobre 2014.
Pdf
- Tahiti.infos (2017) Petite fourmi de feu : est-il déjà trop
tard ? tahiti.infos, 26 juillet 2017, p. http://www.tahiti-infos.com/Petite-fourmi-de-feu-est-il-deja-trop-tard_a163040.html.
Pdf
-
Thiberge, C. (2021) Les invasions biologiques coûtent cher à l’humanité.
lemonde.fr
31 mars 2021.
-
Trigos-Peral, G., S. Abril and E. Angulo (2020). Behavioral responses to numerical
differences when two invasive ants meet: the case of Lasius neglectus and Linepithema
humile. Biological Invasions. doi: 10.1007/s10530-020-02412-4.(Pdf
libre de droits)
- Von Aesch, L.
and D. Cherix (2001). Recrutement alimentaire et recrutement exploratoire chez
la fourmi fantôme Tapinoma melanocephalum : une seule et même stratégie
? Actes Coll. Insectes Soc. 14: 7-11. Pdf
-
Verdo, Y. (2019) Espèces envahissantes : la traque
s'affine. lesechos.fr, 14 septembre 2018, p. https://www.lesechos.fr/idees-debats/sciences-prospective/especes-envahissantes-la-traque-saffine-1131653
. Pdf
- Wohllenben, P. (2017).
La vie
secrète des arbres, Les Arènes. 268p.
Autres références
- Achury,
R., P. Chacón de Ulloa, Á. Arcila and A. V. Suarez (2020). Habitat
disturbance modifies dominance, coexistence, and competitive interactions in
tropical ant communities. Ecological Entomology 45: 1247-1262. doi: 10.1111/een.12908.
- Albouy, V. (2017). Étonnants
envahisseurs Quae. 160p.
- Barbieri, R. F., P.
J. Lester, A. S. Miller and K. G. Ryan (2013). A neurotoxic pesticide changes
the outcome of aggressive interactions between native and invasive ants. Proc
R Soc B 280: 20132157.
- Bates,
O. K., S. Ollier and C. Bertelsmeier (2020). Smaller climatic niche shifts in
invasive than non-invasive alien ant species. Nature Communications 11(1): 5213.
doi: 10.1038/s41467-020-19031-1.
- Bertelsmeier, C. and F. Courchamp (2014). Future ant invasions
in France. Environmental Conservation 41(2): 217-228. 10.1017/s0376892913000556
- Bertelsmeier, C., S. Ollier, A. Avril, O. Blight, H. Jourdan and F. Courchamp
(2016). Colony–colony interactions between highly invasive ants. Basic
and Applied Ecology 17(2): 106-114. http://dx.doi.org/10.1016/j.baae.2015.09.005
- Bertelsmeier, C., S. Ollier, A. Liebhold and L. Keller (2017). Recent human
history governs global ant invasion dynamics. 1: 0184. 10.1038/s41559-017-0184
http://dharmasastra.live.cf.private.-
Arnan, X., E. Angulo, R. Boulay, R. Molowny-Horas, X. Cerdá and J. Retana
(2021). Introduced ant species occupy empty climatic niches in Europe. Scientific
Reports 11(1): 3280. doi: 10.1038/s41598-021-82982-y
springer.com/articles/s41559-017-0184#supplementary-information.
Libre de droits
- Bertelsmeier, C., S. Ollier, A. M. Liebhold, E. G. Brockerhoff, D. Ward and
L. Keller (2018). Recurrent bridgehead effects accelerate global alien ant spread.
Proceedings of the National Academy of Sciences. 10.1073/pnas.1801990115
- Bertelsmeier, C., S. Ollier, A. Liebhold and L. Keller (2017). Recent human
history governs global ant invasion dynamics. Nature Ecology & Evolution
1: 0184. doi: https://doi.org/10.1038/s41559-017-0184.
- Blatrix, R., T. Colin, P. Wegnez, C. Galkowski and P. Geniez (2018). Introduced
ants (Hymenoptera: Formicidae) of mainland France and Belgium, with a focus
on greenhouses. Ann. Soc. Entomol. France (N.S.) 54(4): 293-308. 10.1080/00379271.2018.1490927
- Blight, O., E. Provost, M. Renucci, A. Tirard and J. Orgeas (2010). A native
ant armed to limit the spread of the Argentine ant. Biological Invasions 12(11):
3785-3793. 10.1007/s10530-010-9770-3
- Blight, O., M. Renucci, A. Tirard, J. Orgeas and E. Provost (2010). A new
colony structure of the invasive Argentine ant (Linepithema humile) in Southern
Europe. Biological Invasions 12: 1491-1497.
- Breed, M. (2016). Two views on invasive ants. Insectes Sociaux: 1-2. 10.1007/s00040-016-0494-y
- Buczkowski, G. (2010). Extreme life history plasticity and the evolution of
invasive characteristics in a native ant. Biological Invasions 12: 3343-3349.
- Buczkowski, G. (2016). The Trojan horse approach for managing invasive ants:
a study with Asian needle ants, Pachycondyla chinensis. Biological
Invasions 18(2): 507-515. 10.1007/s10530-015-1023-z
- Campolo, O., V. Palmeri, A. Malacrinò, F. Laudani, C. Castracani, A.
Mori and D. A. Grasso (2015). Interaction between ants and the Mediterranean
fruit fly: New insights for biological control. Biological Control 90(0): 120-127.
http://dx.doi.org/10.1016/j.biocontrol.2015.06.004
- Calcaterra, L. A., A. Cabrera and J. A. Briano (2016). Local co-occurrence
of several highly invasive ants in their native range: are they all ecologically
dominant species? Insectes Soc in press. doi:10.1007/s00040-016-0481-3
- Carson, R. (1968). Le printemps silencieux (Silent Spring, 1962), Livre de
Poche.
- Cerdá, X., E. Angulo, S. Caut and F. Courchamp (2012). Ant community
structure on a small Pacific island: only one native species living with the
invaders. Biological Invasions 14(2): 323-339. 10.1007/s10530-011-0065-0
- Charrier,
N. P., C. Hervet, C. Bonsergent, M. Charrier, L. Malandrin, B. Kaufmann and
J. M. W. Gippet (2020). Invasive in the North: new latitudinal record for Argentine
ants in Europe. Insectes Sociaux 67(2): 331-335. doi: 10.1007/s00040-020-00762-9.
- Cooling, M., S. Hartley, D. A. Sim and P. J. Lester (2012). The widespread
collapse of an invasive species: Argentine ants (Linepithema humile) in New
Zealand. Biology Letters 8: 430-433. DOI: 10.1098/rsbl.2011.1014.
- Cooling, M. and B. D. Hoffmann (2015). Here today, gone tomorrow: declines
and local extinctions of invasive ant populations in the absence of intervention.
Biological Invasions 17(12): 3351-3357. 10.1007/s10530-015-0963-7
- Cooling, M., M. A. M. Gruber, B. D. Hoffmann, A. Sébastien and P. J.
Lester (2016). A metatranscriptomic survey of the invasive yellow crazy ant,
Anoplolepis gracilipes, identifies several potential viral and bacterial pathogens
and mutualists. Insectes Sociaux: 1-11. 10.1007/s00040-016-0531-x
- Cordonnier, M., A. Bellec, G. Escarguel and B. Kaufmann (2020). Effects of
urbanization–climate interactions on range expansion in the invasive European
pavement ant. Basic and Applied Ecology 44: 46-54. https://doi.org/10.1016/j.baae.2020.02.003.
- Courchamp, F.
(2021). "Chaque espèce invasive suit sa niche climatique".
La Recherche Biodiversité et Climat. n°567. 39-40
- Cremer, S., L. V. Ugelvig, F. P. Drijfhout, B. C. Schlick-Steiner,
F. M. Steiner, B. Seifert, D. P. Hughes, A. Schulz, K. S. Petersen, H. Konrad,
et al. (2008). The Evolution of Invasiveness in Garden Ants. PLoS ONE 3(12):
e3838. 10.1371/journal.pone.0003838. libre
de droits
- Darrouzet,
E., J. Gévar, Q. Guignard and S. Aron (2015). Production of early diploid
males by European colonies of the invasive hornet Vespa velutina nigrithorax.
PLOS ONE 10(9): 1-9.
- Dedeine, F., S. Bankhead-Dronnet, C. Lopez-Vaamonde and C. Bouget (2017).
Invasions biologiques : le rôle des microorganismes symbiotiques. Microscoop
HS, octobre 2017: 16-17.
- Dejean, A., R. Céréghino, M. Leponce, V. Rossi, O. Roux, A.
Compin, J. H. C. Delabie and B. Corbara (2015). The fire ant Solenopsis saevissima
and habitat disturbance alter ant communities. Biological Conservation 187(0):
145-153. http://dx.doi.org/10.1016/j.biocon.2015.04.012
- Diagne,
C., B. Leroy, A.-C. Vaissière, R. E. Gozlan, D. Roiz, I. Jaric, J.-M.
Salles, C. J. A. Bradshaw and F. Courchamp (2021). High and rising economic
costs of biological invasions worldwide. Nature. doi: 10.1038/s41586-021-03405-6.
- Dieng, H., R. B. M. Zawawi, N. I. S. B. M. Yusof, A. H. Ahmad, F. Abang, I.
A. Ghani, T. Satho, H. Ahmad, W. F. Zuharah, A. H. A. Majid, et al. (2016).
Green tea and its waste attract workers of formicine ants and kill their workers—implications
for pest management. Industrial Crops and Products 89: 157-166. http://dx.doi.org/10.1016/j.indcrop.2016.05.019
- Eritja, R., J. R. B. Palmer, D. Roiz, I. Sanpera-Calbet and F. Bartumeus (2017).
Direct Evidence of Adult Aedes albopictus Dispersal by Car. Scientific Reports
7(1): 14399. 10.1038/s41598-017-12652-5. libre
de droits
- Espadaler, X. and V. Bernal (2015) Lasius neglectus, a polygynous, sometimes
invasive, ant. 2015, p. http://www.creaf.uab.es/xeg/Lasius/Ingles/index.htm
- Espadaler, X. & Rey, S. Biological constraints and colony founding
in the polygynous invasive ant Lasius neglectus(Hymenoptera, Formicidae). Insectes
Sociaux 48, 159-164 (2001).
- Foucaud, J., J. Orivel, A. Loiseau, J. H. C. Delabie, H. Jourdan, D. Konghouleux,
M. Vonshak, M. Tindo, J.-L. Mercier, D. Fresneau, et al. (2010). Worldwide invasion
by the little fire ant: routes of introduction and eco-evolutionary pathways.
Evolutionary Applications 3(4): 363-374.
- Fournier, D., A. Estoup, J. Orivel, J. Foucaud, H. Jourdan, J. Le Breton and
L. Keller (2005). Clonal reproduction by males and females in the little fire
ant. Nature 435: 1230-1234.
-
Fournier, A., C. Penone, M. G. Pennino and F. Courchamp (2019). Predicting future
invaders and future invasions. Proceedings of the National Academy of Sciences:
201803456. 10.1073/pnas.1803456116
- Frizzi, F., V. Bartalesi and G. Santi (2017). Combined effects of temperature
and interspecific competition on the mortality of the invasive garden ant, Lasius
neglectus: A laboratory study. Journal of Thermal Biology 65: 76-81.
- Garnas, J. R. (2007). Intercolony Aggression Within and Among Local Populations
of the Invasive Ant, Myrmica rubra (Hymenoptera: Formicidae), in Coastal Maine.
Environmental Entomology 36: 105-113.
- Gévar, J., A.-G. Bagnères, J.-P. Christidès and E. Darrouzet
(2017). Chemical Heterogeneity in Inbred European Population of the Invasive
Hornet Vespa velutina nigrithorax. Journal of Chemical Ecology 43:
763–777. 10.1007/s10886-017-0874-4
- Gippet, J. M. W., N. Mondy, J. Diallo-Dudek, A. Bellec, A. Dumet, L. Mistler
and B. Kaufmann (2017). I’m not like everybody else: urbanization factors
shaping spatial distribution of native and invasive ants are species-specific.
Urban Ecosystems 20(1): 157-169. 10.1007/s11252-016-0576-7
- Gippet,
J. M. W., T. Colin, J. Grangier, F. Winkler, M. Haond, A. Dumet, S. Tragust,
N. Mondy and B. Kaufmann (2021). Land-cover and climate factors contribute to
the prevalence of the ectoparasitic fungus Laboulbenia formicarum in its invasive
ant host Lasius neglectus. Fungal Ecology 51: 101045. doi: https://doi.org/10.1016/j.funeco.2021.101045.
- Gippet,
J. M. W. and C. Bertelsmeier (2021). Invasiveness is linked to greater commercial
success in the global pet trade. Proceedings of the National Academy of Sciences
118(14): e2016337118. doi: 10.1073/pnas.2016337118.
- Giraud, T., J. S. Pedersen and L. Keller (2002). Evolution
of supercolonies: the argentine ants of southern Europe. Proc. Natl. Acad. Sci.
U.S.A. 99: 6075-6079.
- Groden, E., F. A. Drummond, J. R. Garnas and A. Francoeur (2005). Distribution
of an Invasive Ant, Myrmica rubra (Hymenoptera: Formicidae), in Maine. J. Econ.
Entomol. 98: 1774_1784.
-
Helanterä, H. (2022). Supercolonies of ants (Hymenoptera: Formicidae):
ecological patterns, behavioural processes and their implications for social
evolution. Myrmecol News 32: 1-22. doi: doi: 10.25849/myrmecol.news_032:001
- Herzberg, N. (2016b). Combattre les invasions biologiques. Le Monde Science
et Médecine Mercredi 6 juillet 2016. p. 1, 4-5.
- Hicks, B. J., B. L. Pilgrim and H. D. Marshall (2014). Origins and genetic
composition of the European fire ant (Hymenoptera: Formicidae) in Newfoundland,
Canada. Canadian entomologist: 1-8.
- Hoffmann, B. D., G. M. Luque, C. Bellard, E. Holmes and C. J. Donlan (2016).
Improving invasive ant eradication as a conservation tool: A review. Biological
Conservation 198: 37-49.
- Jacquot, M., P. Tixier, O. Flores, D. Muru, F. Massol, B. Derepas, F. Chiroleu
and J.-P. Deguine Contrasting predation services of predator and omnivore diversity
mediated by invasive ants in a tropical agroecosystem. Basic and Applied Ecology.
http://dx.doi.org/10.1016/j.baae.2016.09.005
- Joignot, F. (2018a). La mondialisation est aussi biologique. Le Monde 24 février
2018. p. 5.
- Joignot, F. (2018b). La grande plastification. Le Monde Idées 21 avril
2018. p. 5.
- Jolivet, P. (1986). Les fourmis et les plantes, Boubée. 254
- Keller, L. and E. Gordon (2006). La vie des fourmis, Odile Jacob. 304 p.
- Kiers, T. E., T. M. Palmer, A. R. Ives, J. F. Bruno and J. L. Bronstein (2010).
Mutualisms in a changing world: an evolutionary perspective. Ecology Letters
13(12): 1459-1474.
- Klotz, J., L. Hansen, R. Pospischil and M. Rust (2008). Urban ants of North
America and Europe. Cornell, Cornell University Press.
- Klotz, J., D. H. Oi, K. M. Vail and D. F. Williams (1996). Laboratory Evaluation
of a Boric Acid Liquid Bait on Colonies of Tapinoma melanocephalum Argentine
Ants and Pharaoh Ants (Hymenoptera: Formicidae). Journal of Economic Entomology
89: 673-677. DOI: http://dx.doi.org/10.1093/jee/89.3.673
-
Lenoir, A. and E. Perdereau (2022). Les effets de l’alimentation sur les
hydrocarbures cuticulaires de la fourmi invasive Tapinoma magnum. Bull. Soc.
Hist. Nat. Toulouse 158: sous presse. Pdf
- Liang, D. and J. Silverman (2000). "You are what you eat": diet
modifies cuticular hydrocarbons and nestmate recognition in the argentine ant,
Linepithema humile. Naturwissenschaften 87: 412-416.
- Lowe, S. M., M. Browne and S. Boudjelas (2000). 100 of the world's worst invasive
alien species. Aliens 12S: 1-12.
- Luo, Y.-P. and N.-T. Chang (2013). Strategies for controlling the ghost ant,
Tapinoma melanocephalum (Hymenoptera: Formicidae) with liquid bait. Journal
of Asia-Pacific Entomology(0). http://dx.doi.org/10.1016/j.aspen.2012.12.001
- Martins, C., R. F. Souza and O. C. Bueno (2012). Presence and distribution
of the endosymbiont Wolbachia among Solenopsis spp. (Hymenoptera: Formicidae)
from Brazil and its evolutionary history. Journal of Invertebrate Pathology(0).
10.1016/j.jip.2012.01.001
- Ndoutoume-Ndong, A. and J.-B. Mikissa (2007). Influence de la présence
de la fourmi Wasmannia auropunctata (Roger 1863) (Hymenoptera : Formicidae)
sur les autres espèces de fourmis dans la réserve de la Lopé
(centre du Gabon). Ann. soc. entomol. Fr. (n.s.) 43: 155-158.
- Ogogol, R., J. P. Egonyu, G. Bwogi, S. Kyamanywa and M. Erbaugh (2017). Interaction
of the predatory ant Pheidole megacephala (Hymenoptera: Formicidae) with the
polyphagus pest Xylosandrus compactus (Coleoptera: Curculionidea). Biological
Control 104: 66-70. http://dx.doi.org/10.1016/j.biocontrol.2016.11.002
- Passera, L. (1994). Characteristics of tramp species. Exotic ants: Biology,
impact, and control of introduced species. D. F. Williams. Boulder, CO, Westview
Press: 23-43.
- Passera, L. (2006). La véritable histoire des fourmis. Paris, Fayard.
304
- Passera, L. and S. Aron (2005). Les fourmis. Comportement, organisation sociale
et évolution. Ottawa, Presses scientifiques du CNRC. 480pp.
- Pearcy, M., M. A. D. Goodisman and L. Keller (2011). Sib mating without inbreeding
in the longhorn crazy ant. Proceedings. Biological Sciences / The Royal Society
278(1718): 2677-2681. 10.1098/rspb.2010.2562
- Peck, S. B., J. Heraty, B. Landry and B. J. Sinclair (1998). Introduced insect
fauna of an oceanic archipelago: the Galápagos Islands, Ecuador. Am.
Entomol. 44: 218-237.
- Porter, S. D. and L. A. Calcaterra (2013). Dispersal and competitive impacts
of a third fire ant decapitating fly (Pseudacteon obtusus) established in North
Central Florida. Biological Control 64: 66-74. 10.1016/j.biocontrol.2012.09.018
- Porter, S. D., R. M. Plowes and C. E. Causton (2018). The fire ant decapitating
fly, Pseudacteon bifidus (Diptera: Phoridae): host specificity and attraction
to potential food items. Florida Entomologist 101: 55-60.
- Putri,
D., M. Yokozawa, T. Yamanaka and A. L. Cronin (2021). Trait plasticity among
invasive populations of the ant Technomyrmex brunneus in Japan. Animals 11:
2702. doi: doi.org/10.3390/ani11092702.
- Qiu, H.-L., L.-H. Lu, M. P. Zalucki and Y.-R. He (2016).
Metarhizium anisopliae infection alters feeding and trophallactic behavior in
the ant Solenopsis invicta. Journal of Invertebrate Pathology 138: 24-29. http://dx.doi.org/10.1016/j.jip.2016.05.005
- Seifert,
B. (2020). Revision of the Plagiolepis schmitzii group with description of Pl.
invadens sp. nov. - new invasive supercolonial species (Hymenoptera: Formicidae).
Dtsch. Entomol. Z. 67: 183-196.
-
Taheri, A., M. Elmahroussi, J.-L. Reyes-López, N. Bennas and J. C. Brito
(2020). Ants invading deserts: Non-native species in arid Moroccan oases. Journal
of Arid Environments: 104122. doi: https://doi.org/10.1016/j.jaridenv.2020.104122.
- Tragust, S., H. Feldhaar, X. Espadaler and J. Pedersen (2015). Rapid increase
of the parasitic fungus Laboulbenia formicarum in supercolonies of the invasive
garden ant Lasius neglectus. Biological Invasions 17(10): 2795-2801. 10.1007/s10530-015-0917-0
- Turbelin,
A. J., B. D. Malamud and R. A. Francis (2017). Mapping the global state of invasive
alien species: patterns of invasion and policy responses. Global Ecology and
Biogeography 26(1): 78-92. doi: https://doi.org/10.1111/geb.12517.
- Ugelvig, L. V., F. P. Drijfhout, D. J. C. Kronauer, J. J. Boomsma, J. S. Pedersen
and S. Cremer (2008). The introduction history of invasive garden ants in Europe:
Integrating genetic, chemical and behavioural approaches. BMC Biology 6:11:
doi:10.1186/1741-7007-1186-1111.
- Ugelvig, L. V. and S. Cremer (2012). Effects of social immunity and unicoloniality
on host-parasite inetractions in invasive insect societies. Functional Ecology
26: 1300-1312.
- Valles, S. M., D. H. Oi and S. D. Porter (2010). Seasonal variation and the
co-occurrence of four pathogens and a group of parasites among monogyne and
polygyne fire ant colonies. Biological Control 54: 342-348.
- Vincent, C. (2016). Peace et Louves. Pour le philosophe Baptiste Morizot,
l'homme doir créer des relations "diplomatiques" avec le loup
plutôt que de le tuer. Le Monde Idées 25 juin 2016. p. 2.
- Vonshak, M., T. Dayan, J. Foucaud, A. Estoup and A. Hefetz (2009). The interplay
between genetic and environmental effects on colony insularity in the clonal
invasive little fire ant Wasmannia auropunctata. Behavioral Ecology and Sociobiology
63: 1667-1677.
- Wetterer, J. (2008). Worldwide spread of the longhorn crazy ant, Paratrechina
longicornis (Hymenoptera: Formicidae) Myrmecol News 11: 137-149.
- Wetterer, J. and S. D. Porter (2003). The little fire ant Wasmannia auropunctata:
distribution, impact and control. Sociobiology 42: 1-41.
- Wetterer, J. K. (2006). Quotation error, citation copying, and ant extinctions
in Madeira. Scientometrics 67: 351-372.
- Wetterer, J. K. and A. G. Radchenko (2011). Worldwide spread of the ruby ant,
Myrmica rubra (Hymenoptera: Formicidae). Myrmecological News 14: 87-96.
- Wetterer, J. K. (2012). "Worldwide spread of Emery's sneaking ant, Cardiocondyla
emeryi (Hymenoptera: Formicidae)." Myrmecol. News 17: 13-20.
- Wetterer, J. K., D. J. C. Kronauer and M. L. Borowiec (2012). Worldwide spread
of Cerapachys biroi (Hymenoptera: Formicidae: Cerapachyinae). Myrmecol. News
17: 1-4.
- Wilson, E. O. (2000). Naturaliste, Bartillat. 422 p.
- Zhu, Y.-G., M. Gillings,
P. Simonet, D. Stekel, S. Banwart and J. Penuelas (2017). Microbial mass movements.
Science 357(6356): 1099-1100. 10.1126/science.aao300
-
Xu, Y., E. L. Vargo, K. Tsuji and R. Wylie (2022). Exotic Ants of the Asia-Pacific:
Invasion, National Response, and Ongoing Needs. Annual Review of Entomology
67(1): null. doi: 10.1146/annurev-ento-060721-085603. Voir Fourmis
invasives.

{kind=link}
{kind=link}
{kind=link}