
Les plantes et les fourmis
Alain Lenoir mis à jour 11-Mar-2025
Les plantes sont souvent associées à des fourmis, depuis des associations facultatives (consommation, habitat) jusqu’aux associations obligatoires que l’on appelle symbiotiques.
Quelques définitions
:
- Mutualisme : Relation qui apporte des bénéfices aux deux partenaires
- Symbiose : Relation mutualiste permanente entre deux organismes
- Myrmécophytes : plantes supérieures qui entretiennent une relation
mutualiste obligatoire avec des fourmis
Ce texte correspond en grande
partie à des conférences grand public sur les relations plantes
- fourmis. Pour
voir toutes les diapos
Voir le livre de Pierre
Jolivet "Les fourmis et les plantes" (1986) et la synthèse
qu'il a fait en 1991.
Voir une belle synthèse par Luc Passera : Les arbres à fourmis.
Futura
Science 4 mars 2012.
Et une synthèse complète : The
Ecology and Evolution of Ant-Plant Interactions, Victor Rico-Gray et Paulo
S. Oliveira, Chicago University Press 2007. Tout y est..
Fourmis et plantes, un amour préhistorique selon Christophe Josset de l'Express (2018). Un énorme travail de Corie Moreau et collaborateurs qui ont comparé les différences d'ADN de deux groupes d'organismes : 1 700 espèces de fourmis et 10 000 catégories de plantes. Les analyses montrent que leur entraide est bien plus ancienne que prévue : ses prémices remontent à l'époque du mésozoïque, c'est-à-dire entre 66 et 250 millions d'années avant notre ère ! "Chez les fourmis qui misent beaucoup sur les plantes, nous avons même établi un ordre d'apparition progressif de leur dépendance : d'abord les prédatrices se nourrissent occasionnellement de végétaux, puis leur régime évolue à force [pour devenir omnivore voire herbivore], avant de s'en faire finalement un nid", détaille Matthew P. Nelsen. Il a fallu attendre quelques dizaines de millions d'années plus tard pour que les plantes s'y adaptent et s'accommodent finalement de leurs visiteurs. Chacun à son rythme, comme dans un vieux couple. "Les fourmis ont commencé par chercher de la nourriture dans les arbres, ont incorporé les plantes à leur alimentation, et à partir de là, ont commencé à vivre dans les arbres", souligne Corie Moreau (Nelsen et al 2018, voir Josset 2018).
Céline Leroy. Les animaux et les plantes peuvent-ils s’entraider ? , theconversation 10 octobre 2023. Article très intéressant sur le mutualisme entre plantes et pollinisateurs, les fourmis qui protègent les plantes.
1)
Associations facultatives
Nids dans les plantes
De nombreux arbres et arbustes et même des petites plantes sont utilisés
par les fourmis pour nidifier : trous dans le tronc, galles, épines,
branches creuses. En régions tropicales des espèces de fourmis
opportunistes construisent de grands nids en carton ou en terre sur les arbres
(attention, ce sont souvent aussi des termites). Elles élèvent
des homoptères producteurs de sève (voir plus loin).
Il s’agit d’associations non spécifiques : une espèce
de fourmis peut s’installer sur n’importe quel végétal
si les conditions sont favorables.
On connaît des espèces avec individus « portiers »
à tête élargie pour boucher l’entrée du nid
(Colobopsis truncatus dans nos régions).
Lasius fuliginosus fréquente chez nous forme un nid en carton
(fibres végétales mâchées comme les guêpes)
dans une cavité d’arbre.
Les fourmis charpentières vivent dans des galeries du bois mort qu’elles
agrandissent. Chez nous il y a Camponotus vagus qui ne fait pas de
dégâts. Ce n’est pas le cas d’autres fourmis charpentières
en Amérique du Nord qui peuvent attaquer des charpentes.
Passiflore
et héliconius
Didier Van Cauwelaert, dans
"Les
émotions cachées des plantes" Plon (2018) :
"Mais il
est un cas de figure encore plus saisissant, c'est celui où la plante
développe une action solidaire au bénéfice d'un de ses
prédateurs, dont elle a su maîtriser les nuisances tout en récupérant
à son profit l'énergie qu'elle a puisée dans son processus
de riposte. C'est la fascinante histoire de la passiflore et du papillon héliconius.
Au sein de leurs cinq cents espèces respectives, depuis des dizaines
de millions d'années, leur numéro de duettistes fonctionne de
la même manière... Premier temps : le lépidoptère
pond ses oeufs sur les plus jeunes feuilles de la plante grimpante, afin que
ses chenilles trouvent en naissant une nourriture encore comestible. Donc, la
passiflore, si elle se laisse faire, va perdre ses nouvelles pousses et ne sera
plus à même de lan-cer ses vrilles à l'assaut des supports
voisins en vue d'épanouir ses fleurs. Alors, pour tromper le papillon,
elle déguise ses jeunes feuilles en leur donnant la forme de celles de
certaines plantes auxquelles elle s'agrippe pour grimper. Des feuilles dont
elle choisit l'apparence en fonction d'un critère invariable : elles
ne sont pas digérables par les chenilles de l'héliconius, qui
le sait. Une fois parvenue au stade de lumière idéale, en haut
de ses tuteurs, la passiflore fabrique ensuite ses vraies feuilles, lesquelles
sécrètent une substance dont raffolent les fourmis. Celles-ci,
dès lors, avec leur redoutable agressivité, vont empêcher
le papillon de venir pondre dans leur assiette. Sauf que... au fil des siècles,
l'héliconius a su percer le stratagème et le contrer. Une de ses
paires de pattes, nous explique Jean-Marie Pelt dans La Raison du plus faible,
est devenue chimiquement sensible aux feuilles de passiflore, qu'il est donc
désormais capable de reconnaître en dépit de leur camouflage.
Comment la plante va-t-elle réagir à cette perte d'incognito ?
En dotant ses feuilles de petites boules jaunes simulant à la perfection
les oeufs de ce papillon. L'arrivant croit donc qu'un de ses congénères
l'a pris de vitesse, et il va pondre ailleurs, pour éviter la surpopulation
qui priverait sa progéniture d'une nourriture suffisante. Ailleurs, c'est-à-dire
sur l'une des feuilles « libres » que lui laisse la passiflore,
prête à sacrifier, semble-t-il, un certain nombre de ses jeunes
pousses dès lors que son développement global n'est plus menacé."
Élevage
de pucerons, cochenilles et autres homoptères.
Les fourmis sont très attirées par le sucre. Une source de sucre
(et d’acides aminés) est le miellat des pucerons.
Ceux-ci piquent l’écorce des végétaux et pompent
la sève. Ils en absorbent beaucoup trop et rejettent l’excédent
(90%) par l’anus ; c’est le miellat. Les pucerons sont protégés
par les fourmis contre les prédateurs comme les coccinelles et certains
parasites. Par exemple, un travail récent a montré que le taux
de super-parasitisme (parasites de parasites) diminue considérablement
en présence des fourmis Lasius
niger (Sanders and van Veen 2010). Les fourmis vont limiter la dispersion
des pucerons en coupant les ailes des formes ailées ou par action chimique
(les secrétions des glandes mandibulaires des fourmis inhibent le développement
des ailés). De même, les formes aptères se déplacent
moins en présence des fourmis (Oliver et al. 2007). Cela permet une plus
grande production de miellat. Les pucerons et autres « bétail »
peuvent être déplacés en hiver ou lors des migrations. Les
fourrageuses se spécialisent dans l’exploitation d’un groupe
de pucerons, on peut ainsi parler de véritables « troupeaux »
avec leurs bergères (Ebbers and Barrows 1980). Il s’agit donc d’un
véritable élevage. Elles sont parfois aussi adaptées à
leurs fourmis et ne rejettent une gouttelette de miellat que lorsque les fourmis
les sollicitent. Certaines espèces de pucerons ne sont élevées
qu’en présence de leur fourmi hôte et sont reconnues comme
telles grâce à leur odeur qui mime celle des fourmis, ce sont des
hydrocarbures de la cuticule. C’est le cas d’un puceron de la tanaisie,
Macrosiphoniella fuscoviride, par Lasius niger (petite fourmi
noire des jardins, très courante) (Völkl et al. 1999). D’autres
espèces ne sont pas mutualistes et sont mangées par les fourmis.
Il n’y a pas que les hydrocarbures qui permettent la reconnaissance :
les Lasius niger sont attirés par des odeur volatiles des bactéries
Staphylococcus xylosus des pucerons (Aphis fabae)
(Fischer et al. 2015).
On considère souvent les fourmis comme néfastes car la plante
est affaiblie par les pucerons trop nombreux. Mais le bilan est plutôt
considéré positif actuellement car il y a prédation de
nombreux insectes. En fait, le système est complexe : si on place des
filets sur les rosiers pour empêcher le développement du puceron
du rosier on diminue le nombre de fourmis.
Les
fourmis rousses dans nos forêts qui forment les grands
dômes d’aiguilles et branchettes élèvent de nombreux
pucerons mais détruisent de nombreux insectes phytophages. Ces fourmis
rousses sont protégées en Suisse. On les a transplantées
en Italie dans les années 1960, mais avec un succès mitigé.
Il y a actuellement un problème, leur population diminue très
fortement, par exemple en Touraine sans que l’on sache pourquoi. Dans
les forêts du Colorado les ours ouvrent les fourmilières d'une
fourmi des bois Formica pour se nourrir des fourmis et des larves,
ce qui tue la colonie. Des arbustes voisins ne sont plus exploités par
les hémiptères protégés par les fourmis et peuvent
alors de développer mieux (Grangier 2015; Grinath et al. 2015).
Récolte et dispersion des graines
La myrmécochorie (du grec
Myrmecos = fourmi et Chor = porter, disséminer) est une méthode
de dispersion des graines particulièrement efficace.
Tout le monde connaît les fourmis moissonneuses qui font de longues pistes
ramenant au nid des graines en région méditerranéenne.
Ces fourmis moissonneuses habitent en région semi-aride : Messor
chez nous dans le sud (ne piquent pas), Pogonomyrmex en Amérique
(piqûre douloureuse). Un cercle autour du nid marque la limite où
sont rejetées les graines non consommées.
Dans nos régions, en forêt, certaines graines ont un élaiosome
(partie pulpeuse riche sucres, protéines et surtout en graisses attractives
pour les fourmis) ; elles sont rapportées au nid (violette, cyclamen,
mélampyre, chélidoine, euphorbe, anémone hépatique..).
L’élaiosome est consommé, puis la graine est rejetée,
ce qui contribue à sa dissémination. On parle de myrmécochorie
: mode de dispersion des graines par les fourmis. Il existe de nombreuses espèces
de graines avec élaiosome, de couleurs et formes très variées
(au moins 11 000 espèces de plantes adaptées pour la myrmécochorie).
En forêt de Compiègne Formica polyctena disperse diverses
graines plus ou moins rapidement, ce sont les graines de chélidoine et
de mélique à fleur qui le sont le plus rapidement. Carex pilulifera,
Hyacinthoides non-scripta, Allium ursinum, Anemone nemorosa et Colchicum
autumnale sont moins rapidement collectées. Les graines de Paris
quadrifolia sont les seules à ne pas être transportées
(Delatte et Chabrerie 2007).
En Afrique du Sud certaines espèces de plantes endémiques à
élaiosome sont dispersées par des fourmis locales. La fourmi
d’Argentine invasive détruit ces fourmis, mais elle ne récolte
pas ces graines donc les plantes sont menacées de disparition. En fait,
la dissémination des graines se fait à courte distance, en moyenne
2 mètres seulement (avec un maximum de 180m quand même !) (Gómez
and Espadaler 2013).
En Australie il y a Rhytidoponera metallica qui disperse les graines
d'arbres pourvues d'un élaiosome, mais un phasme entoure ses oeufs d'une
substance mimant un élaiosome, les oeufs sont alors ramenés au
nid où ils vons se développer (Kerner 2018, p. 65).
Voir
la myrmécochorie en
Nouvelle-Calédonie, dans la thèse de Le
Yannou-Cateine.
Lubbock en 1883 observait que des fourmis Lasius niger rapportent dans
leur nid des graines de violette "mais j'ignore dans quel but".
Selon Mille
milliards de fourmis : 
Rôle
des fourmis dans la pollinisation ?
Il est très faible, pourtant les fourmis peuvent se nourrir du nectar
des fleurs. Seulement quelques rares cas sont connus (16 cas avérés,
par exemple Camponotus cruentatus avec les euphorbes). Il est possible
que la pollinisation soit empêchée par les substances antiseptiques
des glandes métapleurales qui pourraient tuer les grains de pollen. Certaines
plantes comme le Cytinus hypocistis sont malgré tout dépendantes
des fourmis pollinisatrices qui viennent récolter le nectar des fleurs
et sont attirées par les odeurs de la fleur (de Vega et al. 2014). Les
fourmis visiteuses transportent aussi des levures qui consomment les sucres
du nectar (transformés en alcool), ce qui diminue la qualité du
nectar et le rend moins attractif pour les fourmis (de Vega and Herrera 2012).
Les plantes dopées par les bourdons. C'est fantastique : quand les fleurs de Brassica rapa (moutarde) sont visitées et pollinisées par des bourdons les plantes se portent mieux et ont une meilleure croissance, des fleurs plus parfumées et elles reflètent mieux les ultraviolets comparées à celle des mêmes plantes visitées par des syrphes peu pollinisateurs. Ces moutardes en présence de syrphes s'adaptent en augmentant l'autopollinisation (Le Monde 2017, article de Gervasi et al 2017).
Fourmis
fileuses tropicales Oecophylla.
Le nid est formé de feuilles « cousues » avec la soie des
larves. Il est composé de nombreux petits nids (+ de 150) sur les arbres
qui sont très défendus, il y a une seule reine. Ces fourmis sont
utilisées en Chine et au Vietnam pour protéger les cultures de
citronniers, en Afrique tropicale pour les noix de cajou, cocotiers et cacaoyers.
Autre fourmi fileuse : Polyrhachis
qui fait des nids avec de la soie des larves et des morceaux végétaux
ou même de la terre. Une espèce est utilisée dans la pharmacopée
chinoise, élevée dans ce pays à grande échelle (des
milliers de fermes) et elle sert aussi d'aliment.
Jardins de fourmis sur les arbres en Amazonie : boule de terre sur laquelle poussent de nombreuses plantes comme des Broméliacées et où vivent souvent plusieurs espèces de fourmis. Ces plantes possèdent des nectaires et des corps nourriciers extra floraux qui attirent les fourmis, et produisent un nectar riche en sucres, acides aminés et nombreuses autres substances. Cela produit un véritable écosystème avec des creux où de l’eau s’accumule et toute une faune aquatique s’y installe. La plante récupère de l’azote provenant des cadavres d’insectes aquatiques. En Guyane les chercheurs étudient par exemple la broméliacée Aechmaea mertensii où vivent deux espèces de fourmis, Camponotus femoratus ou Pachycondyla goeldi. La plante change de phénotype selon la fourmi associée (Corbara 2011). Céline Leroy, Chargée de Recherches IRD à ECOFOG, Kourou, travaille sur les associations fongiques dans les racines des broméliacées et sur la transmission du microbiote (bactéries et champignons) des plantes mères vers leur descendance via les graines et plus récemment sur l'influence des fourmis sur les associations champignons / racines (Leroy et al 2022) : "Les jardins de fourmis sont reconnus comme l’un des mutualismes les plus sophistiqués entre des fourmis et des plantes à fleurs, et constituent de bons modèles pour étudier les interactions inter-règnes.Nous avons évalué si les deux espèces de fourmis initiant les jardins de fourmis, Camponotus femoratus et Neoponera goeldii, modifiaient les propriétés physico-chimiques de leurs nids en carton et affectaient la composition et les groupes fonctionnels des communautés fongiques dans les racinaires de la broméliacée Aechmea mertensii. La structure et la composition chimique du nid en carton et la diversité floristique des jardins de fourmis étaient différentes selon l’espèce de fourmis. La diversité et la composition des communautés fongiques racinaires dépendaient de l’identité de la fourmi alors que la diversité fonctionnelle était inchangée. De multiples facteurs biotiques et abiotiques peuvent donc modifier les communautés fongiques associées aux racines. Comme les bénéfices de l’interaction plantes-fourmis dépendent de l’espèce de fourmis, et comme les plantes sont également impliquées dans des interactions avec des endophytes racinaires, cette étude met en évidence l’importance des interactions inter-règnes dans la structuration des communautés fongiques racinaires."
Photo de
Céline Leroy : 

Dans Gérard et les fourmis on parle des fourmis aériennes :

et de fourmis fabriquant du carton :

Selon Des
insectes en société : 
Fourmis
consommatrices de plantes
Certaines fourmis consomment des pétales de fleurs comme les cystes.
Une espèce endémique d’Andalousie est même spécialisée
et a pour cette raison été appelée Cataglyphis floricola.
Fourmis et peupleraies dans la vallée de la Garonne. Les interactions entre jeunes pousses de peuplier en serre et la fourmi Lasius niger (Corenblit et al 2023).
2) Associations obligatoires (= symbioses) entre plantes et fourmis
Plantes
à fourmis (myrmécophytes)
Certaines plantes entretiennent des relations intimes avec les fourmis et forment
une véritable symbiose : hébergement des fourmis dans des cavités
foliaires ou caulinaires (appelées domaties) et nutrition à partir
de corpuscules (corps nourriciers – food bodies- riches en protéines)
ou de secrétions (nectar extrafloral riche en sucres fructose, glucose
et saccharose avec des acides aminés libres), parfois les deux. Dans
l’autre sens les fourmis protègent la plante contre les défoliateurs.
La fourmi apporte aussi des déchets azotés (cadavres des proies)
qui manquent à la plante (comme les plantes carnivores). Pour cette raison,
ces plantes importées et élevées en serre sans leurs fourmis
poussent mal. Sur ces arbres, les fourmis élèvent aussi souvent
dans les cavités des pucerons et autres hémiptères pour
leur miellat (voir plus haut). Les fourmis défendent leur plante hôte
contre les autres plantes comme les nombreuses lianes en forêt tropicale
: elles patrouillent à la base de l’arbuste et coupent les jeunes
pousses. Elles nettoient les feuilles pour éliminer les pathogènes
(champignons – voir plus loin), mangent les œufs des insectes herbivores
et repoussent les mammifères. La plante au début de son développement
produira peu de récompenses pour empêcher la croissance trop rapide
de la colonie de fourmis. Les fourmis sont aussi capables parfois de mutiler
leur plante-hôte pour l’empêcher de fleurir et produire plus
de corps nourriciers. Mais la plante peut se venger de cette castration et produit
des poches foliaires (nids des fourmis) plus petits et moins de corps nourriciers
ce qui limite le développement de la colonie de fourmis. Ainsi s’établit
un équilibre entre la plante et ses hôtes (Fay 2013; Malé
et al. 2013). Voir revue de (Quintero et al. 2013) et Dejean et al (2007) sur
les fourmis de la canopée tropicale.
Autres exemples : les couilles du diable en Asie et îles d'Australie, les fourmis Azteca en Amazonie.
Les plantes à fourmis sont souvent présentes dans les serres tropicales comme au jardin botanique de Nancy. Hélas, les fourmis associées ne sont pas présentes. Si on demande aux responsabless ils disent ne pas vouloir d'insectes qui risquent d'être invasifs... No comment. Pourtant à Nancy on trouve la fourmi Tapinoma melanocephalum, hyper-invasive.
De très nombreux cas de mutualisme plante-fourmis sont connus en zone intertropicale : plus de 100 genres d’Angiospermes et 40 genres de fourmis. Dans nos régions cela n’existe pas.
Divers organismes peuvent profiter du sytème comme des mouches prédatrices des fourmis. Voir Pièges de mouches
Les jardins de fourmis selon
Fred et Jami (C'est
pas sorcier - Les fourmis) : 
Selon Gérard
et les fourmis de Paul Reboux (Flammarion 1932) : 
Medecine man. Film de John Mc Tiernan (1992) met en scène un botaniste qui cherche à extraire un médicament contre le cancer d'une plante de la forêt tropicale. La molécule n'est en fait pas produite par la plante mais par les fourmis qui vivent en association avec elle.
Jardins
du diable
En Amazonie on connaît des surfaces où vit une seule espèce
d’arbuste (Duroia hirsuta) avec sa fourmi (Myrmelachista
schumanni) qui détruit les plantules des autres plantes avec l’acide
formique. La fourmi mord la plantule et y dépose une goutte d’acide
provoquant la nécrose de la feuille en quelques heures. Ces jardins sont
très stables, on en connaît qui ont 800 ans et 350 arbustes. Il
y a une seule colonie avec trois millions d’ouvrières et 15 000
reines (Frederickson et al. 2005). Pour
en savoir plus
Arbre
à la femme adultère
Les arbres Barteria en forêt tropicale hébergent Tetraponera
une fourmi à piqûre très douloureuse. Les femmes adultères
étaient autrefois attachées sur cet arbre au Cameroun. Pour
en savoir plus
Les
arbres acacias
sont myrmécophytes. Ils hébergent des Crematogaster (Afrique)
ou des Pseudomyrmex (Amérique) qui les protègent contre
les défoliateurs, par exemple girafes et éléphants en Afrique.
Des études ont été conduites au Kenya où le nombre
d’éléphants a cru de 5 fois depuis 1992 : quand les fourmis
sont expulsées avec de la fumée, les acacias sont mangés
par les éléphants et un peu par les girafes. La symbiose fourmis
- plantes est donc un élément important dans le maintien de la
savane (Anonyme 2010; Goheen and Palmer 2010).
D’autres Pseudomyrmex
comme P. gracilis sont parasites du système et colonisent l’acacia
sans le défendre.
La flore bactérienne
des feuilles change significativement selon le type de Pseudomyrmex
ou si les mutualistes sont artificiellement enlevées. Dans ce cas les
feuilles sont plus attaquées par des pathogènes comme des champignons.
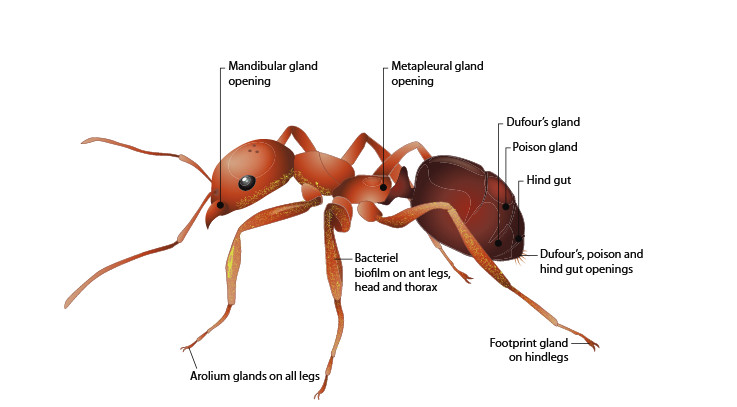
Les bactéries des pattes des fourmis semblent jouer un rôle indirect
dans cette protection (González-Teuber et al. 2014).
Cecropia et Azteca en Amérique : invention du velcro. En Guyane, la symbiose entre les fourmis du genre Azteca et l'arbre Cecropia (ou bois canon, ou parasolier) est fréquente. Les fourmis logent dans les entre-nœuds de la plante. Les chambres ont des fonctions déterminées : pouponnière, réserve de nourriture, salle de repos, décharge.
La fourmi utilise le principe
du Velcro pour s'agripper fermement aux feuilles du Cecropia et pouvoir
ainsi capturer de très grosses proies. Les fourmis de cette espèce
ne se nourrissent pas à partir de corps nourriciers fournis par l'arbre
en plus du logement, mais ont mis au point une stratégie de chasse reposant
sur une organisation sociale très élaborée. Les ouvrières
se postent côte à côte sous la bordure des feuilles de l'arbre
et attendent dans cette position d'éventuelles proies qui viendraient
se poser pour trouver un abri ou attaquer les feuilles de l'arbre. Dans cette
position, les fourmis s'agrippent solidement aux feuilles grâce au principe
du Velcro. En effet, la face inférieure des feuilles présente
une ramification de longs poils qui constitue la partie «velours»
sur laquelle s'accrochent les griffes des ouvrières formant la partie
«crochets». Grâce à ce principe, une fourmi peut maintenir
jusqu'à plus de 5 000 fois son poids. Un groupe d'ouvrières peut
capturer de très grosses proies, la plus grosse rencontrée étant
un criquet de 18,6 g soit 13 350 fois le poids d'une ouvrière (Dejean
et al. 2010). Les Azteca sont
aussi capables de capturer les proies sur leur nid comme les Allomerus.
On les appelle fourmis
tortionnaires.
Les fourmis Azteca qui vivent en symbiose avec les Cecropia
(parasoliers) .
Chaque chambre de la domatie a un microbiome particulier et différent de celui du milieu environnant et les Azteca maintiennent une propreté excellente, surtout autour du couvain. En revanche, elles ne s’occupent pas des agents de maladies pouvant affecter le parasolier ; de ce point de vue, elles ne le protègent pas (Lucas et al 2019). Voir Une leçon de propreté.
Céline Leroy. Les animaux et les plantes peuvent-ils s’entraider ?, theconversation 10 octobre 2023. Article très intéressant sur le mutualisme entre plantes et pollinisateurs, les fourmis qui protègent les plantes.
Le Cecropia, plante
à fourmis : 
Pièges
à insectes (Hirtella / Allomerus)
En Guyane les chercheurs ont beaucoup étudié un arbuste myrmécophyte
: Hirtella physophora qui ne dépasse pas deux mètres
en sous-bois. Les domaties à la base des feuilles hébergent une
petite fourmi prédatrice Allomerus decemarticulata. Elles creusent
des galeries dans les pétioles et les tiges et percent de très
nombreux trous où elles peuvent entrer et sortir. Les trous sont bâtis
avec les poils de la plante, des restes de proies et le système est renforcé
avec le mycélium d’un champignon spécifique Trimmatostroma
cordae. Cela fonctionne comme un piège : les fourmis guettant avec
la tête juste dans le trou et s’agrippant à la proie qui
se pose, pour la piquer (Dejean et al. 2005 - voir fourmis
tortionnaires). On a montré que le champignon est spécifique
et même d’un seul haplotype (ordre des Chaetothyriales) (Ruiz-González
et al. 2011). Le champignon permet aussi le transfert d’azote vers la
plante. C’est une symbiose tripartite complexe (Corbara 2011).
Fourmis
champignonnistes (fourmis parasol en Guadeloupe)
Ce texte est inspiré du livre sur les fourmis coupeuses de feuilles de
(Hölldobler and Wilson 2010) et de sa traduction en français (Hölldobler
and Wilson 2012)
Les coupeuses de feuilles d’Amérique tropicale font partie de la
sous-famille des Attines avec 230 espèces. Elles ont inventé il
y a 50-60 millions d’années la culture des champignons sur compost
élaboré à partir de feuilles mâchées.
Atta et Acromyrmex cultivent un champignon de type lépiote
avec des morceaux de feuilles, les larves sont nourries avec les mycotêtes
(= gondylidia ou choux-raves) du champignon (surtout hémicellulose, la
cellulose est peu digérée), les adultes se nourrissent simplement
avec la sève des feuilles. Le champignon ne fructifie presque jamais,
sauf si la colonie meurt.
Les nids comportent une seule reine inséminée jusqu’à
10 fois. Elle peut vivre jusqu’à 15 ans, elle pond en moyenne 20
œufs par minute, 10 millions par an.
Les vols nuptiaux sont spectaculaires, regroupant des milliers de sexués
(fourmis volantes). La reine fécondée souvent par plusieurs mâles
va avoir une réserve de sperme pour toute sa vie. Elle emporte dans son
vol nuptial un fragment de mycélium pour fonder une nouvelle colonie
et démarrer sa culture.
Le succès considérable de ces fourmis est lié à
la taille des colonies : plusieurs millions d’individus. Ceux-ci sont
de taille très variable (castes), et ont des tâches bien différenciées.
Celles que l’on voit sont les fourrageuses qui vont récolter les
feuilles. Elles s’organisent en brigades et travaillent à la chaîne.
Elles sont protégées par des soldats aux mandibules acérées.
La fourmi et le champignon forment une véritable symbiose. Les ouvrières
reconnaissent leur propre souche de champignon et tout champignon étranger
est rejeté. Il y a un véritable apprentissage de l’odeur
de leur cultivar (Seal et al. 2012). Elles perçoivent même si les
feuilles sont nocives pour le champignon (par exemple traitées avec un
fongicide) et vont alors les éviter (Arenas and Roces 2016). Ces dernières
années on a découvert que la symbiose est bien plus complexe que
ce que l’on pensait. Des champignons parasites très virulents du
genre Escovopsis peuvent se développer dans la culture et tuer
la colonie rapidement. La réponse des fourmis a été de
domestiquer des bactéries du genre Pseudonocardia qui se trouvent
sur le corps de la fourmi et secrètent des antibiotiques. Ces bactéries
sont aussi emportées par la jeune reine fondatrice. Le système
est encore bien plus complexe avec des levures qui mangent les bonnes bactéries
; et d’autres micro-organismes en cours de découverte. On vient
par exemple de trouver des bactéries fixatrices d’azote comme celles
qu’on trouve dans les racines de légumineuses. Il existe un véritable
« microbiome » bactérien dans la meule à champignon
où ce sont des bactéries qui digèrent les parois cellulaires
des plantes (Suen et al. 2010).
Le nid souterrain est de la taille d’une maison. C’est une vraie
mégapole, il faut excaver 60 tonnes pour un nid complet âgé
de seulement 6 ans. Il se prolonge par de nombreux tunnels à 40-50 cm
sous la surface du sol, qui peuvent aller jusqu’à 90 mètres
et facilitent l’approche des arbres ou arbustes. Il peut s’étendre
sur un hectare. Voir une vidéo.
Les fourmis champignonnistes ont aussi une flore bactérienne importante
dans leur estomac, mais elle est simple. Acromyrmex possède
principalement seulement 4 taxa bactériens (Wolbachia, Rhizobiales et
2 Entoplasmatales). Les Rhizobiales sont uniquement extracellulaires dans l’intestin
où elles forment des biofilms (ce qui les protège des antibiotiques
comme la tétracycline) et possèdent des protéines permettant
la fixation d’azote (Sapountzis et al. 2015).
L’agriculture
chez les animaux
L’agriculture n’est pas l’exclusivité des fourmis,
on a vu que certains termites aussi cultivent des champignons. On connaît
d’autres rares exemples comme les coléoptères scolytes qui
creusent un trou dans l’arbre à coloniser et cultivent un champignon
du genre Ambrosia dans les galeries. Les adultes et les larves se nourrissent
du mycélium (Farrellab et al. 2001). Des abeilles sociales trigones ont
aussi domestiqué des champignons qui vivent dans les alvéoles
où ils sont consommés par les larves (Menezes et al. 2015). Très
récemment, on vient de découvrir que les amibes sociales Dictyostelium
discoideum pratiquent une forme primitive d’agriculture : ces amibes
se nourrissent de bactéries et quand la nourriture vient à manquer
ou le milieu devient hostile, elles forment une fructification de spores agrégées.
Certaines souches vont économiser les bactéries qu’elles
consomment et vont les stocker dans leurs fructifications en vue de dispersion
pour le cas où le milieu ne contiendrait pas assez de ces bactéries.
Cependant, seules 30% des amibes vont faire ces réserves ; on ne sait
pas pourquoi (Farrellab et al. 2001; Boomsma 2011; Brillaud 2011; Brock et al.
2011). Ces amibes ont aussi domestiqué une autre souche de bactérie
qui les protège contre les champignons infectieux (Etienne 2013).
Bactéries
et fourmis
On a vu que les champignonnistes sont associées à des bactéries.
De nombreuses fourmis ont des bactéries symbiotes comme les Camponotus.
Celles-ci les aide à assimiler des acides aminés, mais aussi facilitent
leur défense immunitaire (de Souza et al. 2009).
On découvre actuellement l’importance des microbes (unicellulaires,
bactéries, virus) dans le monde vivant : ils jouent un rôle fondamental
dans les écosystèmes. Par exemple, chez l’homme il y a deux
kilogrammes de bactéries dans le tube digestif et l’équilibre
de cette microflore est une garantie de santé. La flore microbienne cutanée
est aussi particulièrement variée. C’est ainsi que certaines
personnes ont une flore plus attractive pour les moustiques anophèles
transmettant le paludisme (Anonyme 2012). En agriculture l’élimination
des pathogènes au bazooka des biocides aboutit à une disparition
du microbiome et de tous ses effets bénéfiques. On connaît
les mycorhizes des plantes supérieures, mais on découvre aussi
qu’il y a des bactéries qui viennent compléter le rôle
des champignons, par exemple en réduisant la consommation d’eau,
ce qui est très important en cas de sécheresse (East 2013). Les
fourmis peuvent aider à la bonne santé des plantes. En effet elles
secrètent des antibiotiques
soit directement soit par leurs bactéries (Voir les glandes
des fourmis). Cela soigne au moins 14 maladies des plantes. On a par exemple
transplanté des fourmis rousses dans une plantation de pommiers, cela
réduit l'apparition de deux maladies (Offenberg et Damgaard 2019; voir
Claudet 2019).
Discussion
Les fourmis sont-elles nuisibles aux plantes ?
Il n’y a pas de réponse simple. La fourmi
de feu est considérée comme très nuisible, en particulier
à cause des piqûres qu’elle inflige, mais dans certaines
conditions elle peut être utile. On a ainsi montré que sur les
plans de coton en serre, que si les aphides sont plus nombreux, ils sont mieux
protégés par la fourmi de feu qui exerce une plus forte prédation
sur les chenilles défoliatrices. Le bénéfice peut être
de 10% en production de coton (Styrsky and Eubanks 2010). Les attines font partie
des écosystèmes tropicaux et ne détruisent pas la forêt
autour de leur nid, mais en monoculture, elles deviennent très nocives.
On a introduit au Gabon une petite fourmi, Wasmannia auropunctata (fourmi
électrique ou petite
fourmi de feu car elle a une piqure très douloureuse) pour protéger
les cacaoyers. Mais par malchance c’est une espèce invasive qui
se répand partout dans le monde tropical… On sait aussi que les
fourmis peuvent transporter des pathogènes des plantes, par exemple dans
les cacaoyères, Pheidole megacephala disperse les champignons
nuisibles du genre Phytophtora.
Il faut considérer les écosystèmes en fonctionnement dynamique
où la biodiversité est fondamentale. On a vu que la biodiversité
favorise le maintien des écosystèmes comme la savane africaine
et cela semble être une loi universelle. D’une manière générale,
elle favorise l’adaptation aux changements et décroît l’émergence
et la transmission de pathogènes comme cela a été montré
sur la tique responsable de la maladie de Lime (Keesing et al. 2010; Le Hir
2010). Voir plus
Plantes
invasives et fourmis
On connaît de nombreuses plantes invasives. On a étudié
les Solidago et on a observé que la richesse en espèces
et en nids de fourmis était plus faible dans les zones envahies (Lenda
et al. 2013).
Notion
de coévolution
C’est une notion très importante. La coévolution désigne
les transformations qui se produisent au cours de l'évolution de deux
espèces vivantes suite à leurs influences réciproques.
Les plantes qui étaient protégées par des fourmis étaient
avantagées et ont été sélectionnées. De même,
les fourmis qui ont su s’adapter à la vie sur ces plantes ont été
sélectionnées au cours de l’évolution. Voir en introduction
l'article de Nelsen et (2018) qui ont fait une étude comparée
de l'ADN de fourmis et plantes.
Communication
entre plantes et fourmis ?
On sait depuis peu que les plantes peuvent communiquer entre elles, par exemple
émettre un signal d’alarme volatil à destination des plantes
voisines quand elles sont attaquées par un herbivore (un insecte le plus
souvent). On vient de montrer que l’arabette des dames (Arabidopsis
thaliana) perçoit les vibrations émises par une chenille
de Pieris rapae en train de manger et qu’elle répond en
émettant du glucosinate et des anthocyanines qui rendent la feuille indigeste.
Si on émet le signal vibratoire la plante réagit de même.
Elle est même capable de différencier ce signal d’un chant
d’insecte ou du bruit du vent, donc de savoir si c’est dangereux
pour elle (Appel and Cocroft 2014).
Quand une plante myrmécophyte est attaquée par un défoliateur,
cela provoque des vibrations qui font sortir les fourmis et les feuilles endommagées
émettent des substances volatiles que les fourmis perçoivent et
qui ont un effet attractif. Il s’agit selon les espèces de terpènes,
aldéhydes, alcools, cétones. Dans un cas, on a trouvé aussi
du méthyle-salicylate (= essence de Wintergreen, odeur de pomme sûre,
très utilisée en parfumerie et arômes alimentaires, précurseur
de l’aspirine, acide salicylique) (Blatrix 2010; Blatrix and Mayer 2010).
La plante semble être capable de détecter la présence des
fourmis. Cela a été montré chez un Piper qui ne
produit des corps nourriciers qu’en présence des fourmis (Blatrix
2010).

Un autre exemple : la plante à tabac. Quand la plante se fait boulotter par une chenille elle dégage une odeur qui attire fourmis et punaises qui mangent la chenille (selon Les insectes en bande dessinée Tome5)

Références
On pourra consulter des ouvrages en français (Keller and Gordon 2006;
Passera 2006; Passera 2008; Hölldobler and Wilson 2012), Voyage chez les
fourmis (Hölldobler and Wilson 1996) et la « bible » des myrmécologues
(Passera and Aron 2005). Pour le côté plantes il existe un excellent
ouvrage qui présente la plupart des cas connus (Jolivet 1986). Pour les
questions de coévolution, biodiversité voir le livre récent
de Pascal Picq (Picq 2013).
Voir
- Blatrix, R. (2010). La communication dans les symbioses entre plantes et fourmis.
Le courrier de la Nature 260: 33-41 Pdf
- de Souza, D., A. Bézier, D. Depoix, J.-M. Drezen and A. Lenoir
(2009). Blochmannia endosymbionts improve colony growth and immune defence in
the ant Camponotus fellah. BMC Microbiology 9(1): 29. Pdf
- Delatte, E. and O. Chabrerie (2007). Dispersion des graines par une espèce
de fourmi rousse des bois (Formica polyctena Foerst.) : approche expérimentale
en forêt de Compiègne (Picardie, France). Symbioses, NS 20: 65-71.
Pdf:
- Jolivet, P. (1991). Les fourmis et les plantes, un exemple de coévolution.
Insectes 83: 3-6. Pdf
-
Josset, C. (2018) Fourmis et plantes, un amour préhistorique.
lexpress.fr, 19 novembre 2018, p. https://www.lexpress.fr/actualite/sciences/fourmis-et-plantes-une-histoire-d-amour-prehistorique_2049228.html.
Pdf
- Kerner, P. (2018). Moi, parasite, Belin. 188p. Pdf
- Le Monde (2017). Les plantes dopées par les bourdons. Le Monde
Science & Médecine Mercredi 22 mars 2017. p. 3. Pdf
- Anonyme (2010). Quand
les fourmis défendent les acacias contre les éléphants.
Le Monde 4 septembre.
- Anonyme (2012). Les moustiques inluencés par l'odeur des humains. Le
Monde 31 décembre 2011- 1-2 janvier 2012. p. 3.
- Appel, H. M. and R. B. Cocroft (2014). Plants respond to leaf vibrations caused
by insect herbivore chewing. Oecologia 175(4): 1257-1266. 10.1007/s00442-014-2995-6
- Arenas, A. and F. Roces (2016). Gardeners and midden workers in leaf-cutting
ants learn to avoid plants unsuitable for the fungus at their worksites. Anim
Behav 115: 167-174.
- Barelli, P. (2011). Vie et mort de l'algue tueuse : la saga de "Caulerpa
taxifolia". Le Monde 4-5 septembre. p. 7.
- Basset, Y., L. Cizek, P. Cuénoud, R. K. Didham, F. Guilhaumon, O. Missa,
V. Novotny, F. Ødegaard, T. Roslin, J. Schmidl, et al. (2012). Arthropod
Diversity in a Tropical Forest. Science 338(6113): 1481-1484. 10.1126/science.1226727
- Blatrix, R. and V. Mayer (2010). Communication in ant-plant symbioses. Plant
communication from an ecological perspective. F. Baluska and V. Ninkivic. Berlin,
Springer: 127-158.
- Boomsma, J. J. (2011). Evolutionary biology: Farming writ small. Nature 469(7330):
308-309.
- Brillaud, R. (2011). Des organismes primitifs pratiquent l'agriculture ! Science
et Vie 1122: p. 13.
- Brock, D. A., T. E. Douglas, D. C. Queller and J. E. Strassmann (2011). Primitive
agriculture in a social amoeba. Nature 469(7330): 393-396.
- Corbara, B. (2011). L'arbuste, les fourmis et le champignon. Espèces
1: 66.
- Corbara, B. (2011). Portrait d'une chercheuse : Céline leroy. Espèces
1.
- de Vega, C. and C. M. Herrera (2012). Relationships among nectar-dwelling
yeasts, flowers and ants: patterns and incidence on nectar traits. Oikos: no-no.
10.1111/j.1600-0706.2012.20295.x
-
Corenblit, D., B. Corbara, R. Cereghino, A. Dejean, P. Duran, V. Garófano-Gómez,
F. Gilbert, E. González-Sargas, F. Julien, L. Lambs, et al. (2023). Effects
of ants on riparian poplars: an ex situ experiment of biotic interaction. Arthropod-Plant
Interactions. 10.1007/s11829-023-09950-3
- de Vega, C., C. M. Herrera and S. Dötterl (2014). Floral volatiles play
a key role in specialized ant pollination. Perspectives in Plant Ecology, Evolution
and Systematics in press(0). http://dx.doi.org/10.1016/j.ppees.2013.11.002
- Dejean, A., C. Leroy, B. Corbara, O. Roux, R. Céréghino, J.
Orivel and R. Boulay (2010). Arboreal ants use the "velcro principle"
to capture very large prey. PLos One 5: e11331.
- Dejean, A., P. J. Solano, J. Ayroles, B. Corbara and J. Orivel (2005). Arboreal
ants build a trap to ambush and capture prey. Nature 434: 793.
- Dejean, A.,
B. Corbara, J. Orivel and M. Leponce (2007). Rainforest canopy ants: the implications
of territoriality and predatory behavior. Functional Ecosystems Communities
1: 105-120
- East, R. (2013). Microbiome: Soil science comes to life. Nature 501(7468):
S18-S19. 10.1038/501S18a
- Ebbers, B. C. and E. M. Barrows (1980). Individual ants specialize on particular
aphids herds (Hymenoptera: Formicidae; Homoptera: Aphididae). Proceeding of
the Entomological Society Washington 82: 405-407.
- Etienne, V. (2013). Pour se protéger, une amibe cultive ses propres
pesticides. Science et Vie Octobre: p. 31.
- European commission DG Env (2015). European salamnders and newts under threat
from fungus by wildlife trade. Science for Environment Policy 402(5 February):
1.
- Farrellab, B. D., A. S. Sequeiraa, B. C. O'Mearaa, B. B. Normarkc, J. H. Chungd
and B. H. Jordale (2001). The evolution of agriculture in beetles (Curculionidas:
Scolytinae nad Platypodinae). Evolution 55: 2011-2027.
- Fay, S. (2013). Une plante "punit" les fourmis trop voraces. Science
et Vie 1153: p. 29.
- Fischer, C., G. Lognay, C. Detrain, M. Heil, A. Grigorescu, A. Sabri, P. Thonart,
E. Haubruge and F. Verheggen (2015). Bacteria may enhance species association
in an ant–aphid mutualistic relationship. Chemoecology: 1-10. 10.1007/s00049-015-0188-3
- Frederickson, M. E., M. J. Greene and D. M. Gordon (2005). 'Devil's gardens'
bedevilled by ants. Nature 437: 495-496.
- Gervasi, D. D. L. and F. P. Schiestl (2017). Real-time divergent evolution
in plants driven by pollinators. Nature Communications 8: 14691. 10.1038/ncomms14691
http://www.nature.com/articles/ncomms14691#supplementary-information. (Libre
de droits)
- Goheen, J. R. and T. M. Palmer (2010). Defensive plant-ants stabilize megaherbivore-driven
landscape change in an African savanna. Current Biology 20: 1-5.
- Gómez, C. and X. Espadaler (2013). An update of the world survey of
myrmechorous dispersal distances. Ecography 36: in press.
- González-Teuber, M., M. Kaltenpoth and W. Boland (2014). Mutualistic
ants as an indirect defence against leaf pathogens. The New Phytologist 202(2):
640-650. 10.1111/nph.12664
- Grangier, J. (2015). Les ours ont la main verte. Espèces 15: 10.
- Grinath, J. B., B. D. Inouye and N. Underwood (2015). Bears benefit plants
via a cascade with both antagonistic and mutualistic interactions. Ecology Letters
18(2): 164-173. 10.1111/ele.12396
- Hölldobler, B. and E. O. Wilson (1996). Voyage chez les fourmis, Le Seuil.
254
- Hölldobler, B. and E. O. Wilson (2010). The leaf-cutting ants. Civilization
by instinct, Norton. 160
- Hölldobler, B. and E. O. Wilson (2012). L'incroyable instinct des fourmis,
Flammarion. 204
- Jacquet, L. (2013). Il était une forêt.
- Jolivet, P. (1986). Les fourmis et les plantes, Boubée. 254
- Joly, J. (2012). Près de 18000 espèces découvertes en
2011. Le Monde 3 janvier. p. 7.
- Keesing, F., L. K. Belden, P. Daszak, A. Dobson, C. D. Harvell, R. D. Holt,
P. Hudson, A. Jolles, K. E. Jones, C. E. Mitchell, et al. (2010). Impacts of
biodiversity on the emergence and transmission of infectious diseases. 468(7324):
647-652.
- Keller, L. and E. Gordon (2006). La vie des fourmis. Paris, Odile Jacob. 304
- Le Hir, P. (2010). Moins d'espèces, plus de maladies infectieuses.
Le Monde 15 décembre. p. 4.
- Lenda, M., M. Witek, P. Skórka, D. Moron and M. Woyciechowski (2013).
Invasive alien plants affect grassland ant communities, colony size and foraging
behaviour. Biological Invasions 15(11): 2403-2414. 10.1007/s10530-013-0461-8
-
Leroy, C., A. Q. Maes, E. Louisanna, J.-F. Carrias, R. Céréghino,
B. Corbara and N. Séjalon-Delmas Ants mediate community composition of
root-associated fungi in an ant-plant mutualism. Biotropica n/a(n/a). doi: https://doi.org/10.1111/btp.13079.
- Lubbock, J. (1883). Fourmis, Abeilles et Guêpes, Librairie Germer Baillère
et Cie. 196p.
- Lucas, J. M., A. A. Madden, C. A. Penick, M. J. Epps, P. R. Marting, J. L.
Stevens, D. J. Fergus, R. R. Dunn and E. K. Meineke (2019). Azteca ants maintain
unique microbiomes across functionally distinct nest chambers. Proceedings of
the Royal Society B: Biological Sciences 286(1908): 20191026. doi:10.1098/rspb.2019.1026.
Libre
de droits
- Malé, P.-J. G., J.-B. Ferdy, C. Leroy, O. Roux, J. Lauth, A. Avilez,
A. Dejean, A. Quilichini and J. Orivel (2013). Retaliation in Response to Castration
Promotes a Low Level of Virulence in an Ant–Plant Mutualism. Evolutionary
Biology: 1-7. 10.1007/s11692-013-9242-7
- Martel, A., M. Blooi, C. Adriaensen, P. Van Rooij, W. Beukema, M. C. Fisher,
R. A. Farrer, B. R. Schmidt, U. Tobler, K. Goka, et al. (2014). Recent introduction
of a chytrid fungus endangers Western Palearctic salamanders. Science 346(6209):
630-631. 10.1126/science.1258268
- Menezes, C., A. Vollet-Neto, A. J. Marsiaoli, D. Zampieri, I. Cardoso Fontura,
A. Ducati Luchessi and V. L. Imperatriz-Fonseca (2015). A brazilian social bee
must cultivate fungus to survive. Current Biology in press. 10.1016/j.cub.2015.09.028
- Mulot, R. (2012). Les sols français en relative bonne santé.
Sciences et Avenir Janvier: p. 28.
- Nelsen,
M. P., R. H. Ree and C. S. Moreau (2018). Ant–plant interactions evolved
through increasing interdependence. Proceedings of the National Academy of Sciences.
10.1073/pnas.1719794115
- Oliver, T. H., A. Mashanova, S. R. Leather, J. M. Cook and V. A. A. Jansen
(2007). Ant semiochemicals limit apterous aphid dispersal. Proceedings of the
Royal Society B: Biological Sciences 274(1629): 3127-3131. 10.1098/rspb.2007.1251
- Passera, L. (2006). La véritable histoire des fourmis. Paris, Fayard.
304
- Passera, L. (2008). Le monde extraordinaire des fourmis. Paris, Fayard.
- Passera, L. and S. Aron (2005). Les fourmis. Comportement, organisation sociale
et évolution. Ottawa, Presses scientifiques du CNRC. 480pp.
Picq, P. (2013). De Darwin à Lévy-Strauss. L'homme et la diversité
en danger. Paris, Odile Jacob. 290
- Quintero, C., K. E. Barton and K. Boege (2013). The ontogeny of plant indirect
defenses. Perspectives in Plant Ecology, Evolution and Systematics(0). http://dx.doi.org/10.1016/j.ppees.2013.08.003
- Ruiz-González, M. X., P.-J. G. Malé, C. Leroy, A. Dejean, H.
Gryta, P. Jargeat, A. Quilichini and J. Orivel (2011). Specific, non-nutritional
association between an ascomycete fungus and Allomerus plant-ants. Biology letters
7(3): 475-479. 10.1098/rsbl.2010.0920
- Sanders, D. and F. J. F. van Veen (2010). The impact of an ant-aphid mutualism
on the functional composition of the secondary parasitoid community. Ecological
Entomology 35(6): 704-710.
- Sapountzis, P., M. Zhukova, L. H. Hansen, S. J. Sorensen, M. Schiott and J.
Boomsma (2015). Acromyrmex Leaf-Cutting Ants Have Simple Gut Microbiota with
Nitrogen-Fixing Potential Appl. Environ. Microbiol. 81: 5527-5537.
- Seal, J. N., J. Gus and U. G. Mueller (2012). Fungus-gardening ants prefer
native fungal species: do ants control their crops? Behavioral Ecology 23(6):
1250-1256. 10.1093/beheco/ars109
- Styrsky, J. D. and M. D. Eubanks (2010). A facultative mutualism between aphids
and an invasive ant increases plant reproduction. Ecological Entomology 35(2):
190-199.
- Suen, G., J. J. Scott, F. O. Aylward, S. M. Adams, S. G. Tringe, A. n. A.
Pinto-Tomás, C. E. Foster, M. Pauly, P. J. Weimer, K. W. Barry,
et al. (2010). An Insect Herbivore Microbiome with High Plant Biomass-Degrading
Capacity. PLoS Genet 6(9): e1001129.
- Vincent, C. (2010). Un sommet pour sauver le tigre de la disparition. Le Monde
21-22 novembre. p. 4.
- Vincent, C. (2011). L'"homme de la forêt" victime de l'huile
de palme. Le Monde 23 décembre. p. 7.
- Vincent, C. (2011). Plus d'une espèce de conifères sur quatre
est menacée dans le monde. Le Monde 11 novembre. p. 10.
- Vincent, C. (2012). Tigre ou ver de terre : qui vaut-il mieux protéger
? Le Monde 5-6 Février. p. 6.
- Völkl, W., J. Woodring, M. Fischer, M. W. Lorenz and K. H. Hoffmann (1999).
Ant-aphid mutualisms: the impact of honeydew production and honeydew sugar composition
on ant preferences. Oecologia, Springer Berlin / Heidelberg. 118: 483-491.
{kind=link}
{kind=link}